

文摘
内皮分化基因(Edg)蛋白质是G-protein-coupled受体激活lysophospholipid介质:sphingosine-1-phosphate (S1P)或lysophosphatidic酸。我们表明,在中枢神经系统,表达Edg8 / S1P5,高亲和性S1P受体,仅限于少突胶质细胞,并表示在发展不成熟阶段的成熟myelin-forming细胞。S1P激活Edg8 / S1P5 O4-positive pre-oligodendrocytes诱导过程收缩通过ρ激酶/ collapsin response-mediated蛋白信号通路,而没有引起收缩在这些细胞S1P源自Edg8 / S1P5-deficient老鼠。Edg8 / S1P5-mediated过程收缩不再局限于未成熟的细胞,观察到后来的发展阶段。相比之下,S1P激活促进成熟的少突胶质细胞的生存而不是pre-oligodendrocytes。S1P-induced生存成熟的少突胶质细胞是介导通过百日咳toxin-sensitive Akt-dependent途径。我们的数据表明,Edg8 / S1P5激活oligodendroglial细胞调节两种截然不同的功能途径调解过程收缩或细胞生存和这些影响取决于细胞的发育阶段。
介绍
少突胶质细胞是中枢神经系统的myelin-forming细胞。轴突的髓鞘形成允许建立一个动作电位通过跳跃式传导的快速传播。在胚胎发育过程中,细胞的少突细胞谱系出现心室区。连续发展阶段(每个被电池的抗体)标识(见补充表1,可用www.jneurosci.org作为补充材料),那么将增殖少突细胞祖转换为一个成熟postmitotic髓鞘少突细胞。这些发展阶段与不同的扩散能力,迁移,分化,生存,成熟,最后髓磷脂沉积,细胞自动还取决于各种外在因素。生长因子(a, FGF-2 IGF-I,据,调节,和神经营养因子)已经被证明能够影响扩散,生存和少突细胞前体细胞的分化(信息公开化)(巴尔et al ., 1992;邦萨尔和普费弗,1997;弗洛雷斯et al ., 2000;Sim et al ., 2002)。指导分子,如semaphorins 3 a, 3 f和netrin-1,指导信息公开化的迁移在开发(杉本et al ., 2001;斯帕斯基et al ., 2002;蔡和米勒,2002年;Jarjour et al ., 2003)。成熟的少突胶质细胞成myelin-forming细胞依赖于生长因子如FGF,据家庭成员(Stankoff et al ., 2002 a;哦,et al ., 2003)。
内皮分化基因(Edg)家庭是由八个G-protein-coupled受体。Sphingosine-1-phosphate (S1P)和/或lysophosphatidic酸(LPA)、相关lysophospholipid介质结构,已被确认为所有已知Edg受体的内源性配体。Edg受体家族可以分成两个子组基于配体特异性和同源性。第一小组结合S1P和由Edg1 / S1P1 Edg3 / S1P3 Edg5 / S1P2 Edg6 / S1P4, Edg8 / S1P5。第二小组结合LPA和包含Edg2 / LPA1 Edg4 / LPA2, Edg7 / LPA3 (李et al ., 1998;范Brocklyn et al ., 1998;春et al ., 1999;Im et al ., 2001)。除了Edg6以外,所有其他Edg受体表达在中枢神经系统。Edg2 / LPA1和Edg8 / S1P5 mRNA表达主要是中枢神经系统白质(Allard et al ., 1998;Im et al ., 2001)。在细胞水平上,Edg2 / LPA1已被证明是成熟的少突胶质细胞表达(Allard et al ., 1998;赛尔维拉et al ., 2002;Stankoff et al ., 2002 b),而它的表情不成熟的少突胶质细胞仍存在争议(穆勒et al ., 1999;道森et al ., 2003)。免疫组织化学分析表明,Edg8 / S1P5表达NG2-positive公开化而不是成熟的少突胶质细胞(Terai et al ., 2003),而表达的Edg8 / S1P5成绩单已经检测到逆转录(RT) pcr在差异化的老鼠少突胶质细胞在文化(Yu et al ., 2004)。
在这里,我们报告Edg8 / S1P5受体已经被公开化,仍然表达了整个开发的少突细胞谱系,包括成熟的髓鞘少突胶质细胞。此外,通过分析对少突细胞S1P文化的影响,我们提供的证据的二价角色Edg8 / S1P5 oligodendroglial血统的受体细胞内信号依赖于它们的分化阶段。S1P-induced Edg8 / S1P5受体的激活O4+pre-oligodendrocytes导致收缩oligodendroglial流程,通过ρkinase-dependent通路包括磷酸化collapsin response-mediated蛋白质(CRMP2)。相反,在成熟的少突胶质细胞,S1P-induced Edg8 / S1P5受体激活导致增加细胞生存,通过Akt介导的信号通路。这些结果表明,Edg8 / S1P5受体信号可以少突细胞发展的特定阶段不同的影响。
材料和方法
动物。可以远系小鼠或Wistar鼠获得Iffacredo (L 'Arbresle、法国)。转基因1900 bp -MBP-lacZ老鼠(麻醉品et al ., 1992),Edg8 / S1P5- / -我们的动物房间里老鼠了。纯合子的Edg8 / S1P5- / -小鼠没有明显行为赤字,和神经病理学的检查Edg8 / S1P5- / -大脑没有任何明显的髓磷脂不足(补充图S1,可用www.jneurosci.org作为补充材料)
目标的Edg8 / S1P5基因和代突变小鼠。基因打靶进行胚胎14.1天的胚胎干细胞(ES)细胞,取代所有的Edg8 / S1P5开放阅读框的下游SrfI限制站点(6英国石油下游的翻译起始密码子)的IRES-lacZ表达盒式磁带包含和积极的选择新霉素磷酸转移酶基因由PGK启动子(图1一个)。5′和3′同源臂(4.1∼kb生态RV / SrfI和∼3.4 kb生态RI /后三世限制片段,分别)从129年克隆svj基因组细菌人工染色体图书馆和放置的两侧IRES-lacZ表达盒式磁带和积极的选择生成目标结构。同源重组neomycin-resistant ES细胞的5′末端目标轨迹由印迹的决定BamHI-digested ES细胞基因组DNA,用437个基点5′外部探测器由PCR(引物:5′-GCTACATTAGAGAGCTCTCCC-3′, 5′-CCTCTTGGTTCTGTTTCTCCC-3′),该检测7.6∼∼4.9 kb野生型和有针对性的轨迹,分别。大约在30 G418-resistant克隆一个经历了同源重组。同源重组的3′末端被确认在这些ES细胞克隆的印迹Xho我领悟了使用243个基点PCR基因组DNA片段的3′外部探测器(引物:5′-CCAGAGGTGGAACTTAGGTGG-3′, 5′-GTGCTCCAGAGACACACTCTC-3′),该检测11.6∼> 16 kb乐队在野生型和有针对性的轨迹,分别。三个定向克隆注入C57BL / 6 j-derived胚泡。男性嵌合体交叉与C57BL / 6 j雌性后代产生N1F0 (C57BL / 6 j×129 ola / Sv),随后是back-crossed四次在C57BL / 6 j背景然后intercrossed产生N5F1代后续测试中使用。N1F0后代pcr证实了上述印迹过程。基因型分析的老鼠N5F1研究人口的生成是由PCR的DNA。引物设计生成特定于野生型的PCR产物轨迹(5′底漆外显子具体,5′-CCAACAGCTTGCAGCGATCCCC-3′;3′底漆外显子具体,5′-GGTTGCTACTCCAGGACTGCCG-3′;30个周期在94°C(30岁),60°C(30岁),和72°C(60)使用,给170个基点的产品尺寸)或目标轨迹(5′,neo gene-specific 5′底漆:5′-CCGGCCGCTTGGGTGGAGAGG-3′;3′,neo gene-specific 3′底漆:5′-TCGGCAGGAGCAAGGTGAGATGACA-3′;30个周期在94°C(30岁),68°C(30岁),和72°C(30岁)被使用,给一个产品299个基点)。 The successful depletion of theEdg8 / S1P5记录证实了rt - pcr进行RNA提取野生型、杂合的,纯合子Edg8 / S1P5突变体的大脑(图1B)。RNA制备使用RNeasy工具包(试剂盒,克劳利,英国)。使用一步法rt - pcr进行rt - pcr试剂盒(试剂盒)使用Edg8 / S1P5特殊引物(5′CCAACAGCTTGCAGCGATCCCC-3′, 5′-GGTTGCTACTCCAGGACTGCCG-3′,产生170个基点的产品),作为一个积极的控制,产生hprt特殊引物(5′-GCTGGTGAAAAGGACCTCT-3′, 5′CACAGGACTAGAACACCTGC-3′,产生250个基点的产品)。PCR条件如下:30年代在94°C, 30年代60°C,在72°C 30和60年代周期。所有实验动物根据英国的要求(科学程序)法案》(1986)和严格符合道德标准。

Edg8 / S1P5定位策略。一个同源重组的示意图表示主要的替代Edg8 / S1P5开放阅读框(ORF)IRES-lacZ,详细的材料和方法。Brt - pcr分析,进行200 ng的总RNA与野生型老鼠的大脑和杂合的和纯合子Edg8 / S1P5无效等位基因。产生HPRT引物被用作控制。
抗体和试剂。抗体被用于以下稀释:anti-myelin碱性蛋白(MBP)单克隆抗体(mAb)(鼠标IgG1 1:50;Euromedex、Souffelweyersheim、法国),anti-glial原纤维酸性蛋白(GFAP)(鼠标马伯,1:200)(雅克et al ., 1986),A2B5马伯(鼠标IgM 1:5;美式文化收藏、马纳萨斯,弗吉尼亚州),O4马伯(鼠标IgM 1:10) (大梁Schachner, 1982),anti-proteolipid蛋白(PLP)(鼠马伯克隆AA3, 1:10) (Yamamura et al ., 1991),anti-β-galactosidase(兔多克隆抗体,1:1000;西切斯特,工具论Technika PA), antigalactosylceramide (GalC)(鼠标马伯IgG3 1:10) (Ranscht et al ., 1982),鼠anti-NG2抗体(1:400)(Diers-Fenger et al ., 2001),antiadenomatous息肉病杆菌(APC)抗体(鼠标马伯,1:50;国际VWR Fontenay苏木香、法国)(Bhat et al ., 1996),anti-MAC1抗体(鼠马伯IgG2b 1:10 0;Ozyme,圣昆汀·雷诺,法国),anti-F4 - 80抗体(鼠马伯IgG2b 1:200;Serotec、Cergy、法国),anti-MAP2抗体(鼠标马伯IgG1, 1:10 0;σ,圣路易斯,密苏里州),SMI31/32抗体(鼠标马伯IgG1 1:1000;斯特恩伯格单克隆抗体,Lutherville博士)。兔子anti-rat Edg8 / S1P5抗血清产生极端的c端12个氨基酸的老鼠Edg8 / S1P5受体序列(CTANRTLVPDATD;序列中的第一个半胱氨酸不但是添加作为一个链接器连接的肽KLH载体免疫)。肽抗体定制了表达载体(CA)圣地亚哥根据标准协议和具有免疫印迹蛋白质提取物正常老鼠的大脑(一个乐队在37 kDa),没有染色Edg8 / S1P5- / -老鼠大脑蛋白质提取(补充图。S2,可用www.jneurosci.org作为补充材料)。采用抗血清稀释在细胞培养和1:1000 1:200组织部分。Alexa(488或594)共轭羊anti-mouse IgG1 IgG3 Alexa-conjugated山羊anti-mouse IgM, anti-rabbit免疫球蛋白,和anti-rat免疫球蛋白来自分子探针(尤金,或者)和在1:1000稀释使用。对于西方墨点法,我们使用以下稀释:anti-phosphorylated Akt(鼠标马伯IgG2b 1:1000;Ozyme), anti-phosphorylated CRMP2抗体(兔多克隆,1:50)(松井et al ., 1996),兔子anti-Edg2 / LPA1抗血清(1:1000)(Stankoff et al ., 2002 b),peroxidase-conjugated anti-mouse和anti-rabbit Ig (1:5000;Dako斯特鲁普、丹麦)。
从σS1P购买。S1P被添加到文化、自由脂肪酸的牛血清白蛋白(σ)使用。百日咳毒素(PTX)从列表获得生物实验室(坎贝尔,CA)和用于最终的浓度200 ng / ml。特定的ρ激酶抑制剂,Y27632 (VWR国际),是用于最终的浓度10μ米。3 Semaphorin条件培养基是来自人类胚胎肾细胞(HEK 293)与某3表达载体,转染如前所述(斯帕斯基et al ., 2002)。
原位杂化。的表达Edg8 / S1P5信使rna可以的产后中枢神经系统或1900年MBP-lacZ分析了老鼠大脑矢状部分原位digoxigenin-labeled小鼠反义riboprobe杂交。老鼠Edg8 / S1P5在pcDNA3.1 V5 His-TOPO subcloned pBluescript II KS + phagemid。部分意义上和反义治疗Edg8 / S1P5cRNA探测器生成的T3和T7启动子和贴上digoxigenin-UTP(勃,曼海姆,德国)。原位杂交和digoxigenin-labeled探针检测进行如前所述(斯帕斯基et al ., 1998)。的特异性染色证明缺乏杂交信号的探测器。
细胞培养。文化进行6 -或24-well板块(合演公司,剑桥,英国)保利-l赖氨酸(PLL)(σ)涂层玻璃盖玻片(OSI, Maurepas,法国)或直接在PLL-coated塑料盘子。少突细胞文化保持在含0.5%胎牛血清的培养基(FCS), 10μ米胰岛素,100μg /毫升转铁蛋白,白蛋白0.28μg /毫升、60 ng / ml黄体酮,16μg腐胺/毫升、40 ng / ml三碘甲状腺氨酸,和30 ng / mll甲状腺素。不同类型的oligodendroglial文化进行:
主要成熟大鼠寡树突胶质细胞(immunolocalization Edg8 / S1P5 S1P对细胞生存的影响的分析和收缩的过程,和生化检测)还是纯种老鼠的大脑被净化后Lubetzki et al。(描述的过程1986年)。阿糖胞苷(105μ米)被添加到培养基中,以避免污染星形胶质细胞的增殖。镀三天后,成熟oligodendroglial人口,评估PLP免疫反应性,代表至少90%的总细胞。
丰富的主要文化O4+/ GalC- - - - - -pre-oligodendrocytes (immunolocalization Edg8 / S1P5 oligodendroglial发展的不同阶段,分析在收缩过程中S1P)的影响是可以获得新生小鼠前脑使用Percoll密度梯度如前所述(Lubetzki et al ., 1991)。镀后三天(即。,at the time when S1P was added to the culture medium), O4+/ GalC- - - - - -pre-oligodendrocytes代表占总细胞数的65%,而6%的细胞O4+/ GalC+不成熟的少突胶质细胞。Edg8 / S1P5-deficient文化源自的大脑Edg8 / S1P5- / -动物,根据相同的技术。
来获得高纯度的文化pre-oligodendrocytes(生存分析和生化实验),oligodendroglial细胞被孤立的“摇晃”后从麦卡锡和De Vellis程序修改(1980年)使用木瓜蛋白酶(30 U /毫升)的分离步骤。金属堆焊两天后,细胞O4的98%+/ GalC- - - - - -pre-oligodendrocytes。
免疫组织化学。双immunolabeling如前所述执行细胞培养(查尔斯et al ., 2000)。对双immunolabeling组织部分,在饱和含10% FCS和50%绵羊血清DMEM 1 h,一夜之间,主要的抗体被孵化PBS中含0.1% Triton x - 100。在PBS洗涤后,Alexa-conjugated二级抗体是孵化在同一介质1 h。Edg8 / S1P5检测,一个附加的步骤微波治疗部分(750 W 0.01的2分钟米柠檬酸缓冲)之前执行孵化与第一抗体。为免疫细胞化学结合原位组织部分杂交,跟着immunolabeling一步原位杂交过程。
(3)- 4 5-Dimethylthiazol-2-yl 2, 5-diphenyltetrazolium溴化生存分析。后3 d培养基,纯化preoligodendrocytes (O4+/ GalC- - - - - -(PLP)或成熟少突细胞+)改为剥夺介质(DM),没有胰岛素和FCS。S1P被添加浓度增加,生存是量化24 h后,由3 - (4 5-dimethylthiazol-2-yl) 2, 5-diphenyltetrazolium溴化(MTT)(σ)测定。结果,表示为一个百分比的控制(即。,细胞在胰岛素(10μ米)和FCS(0.5%)补充中,均值±SEM的至少三个独立的实验中,至少有三个文化/条件。使用费舍尔PSLD测试执行统计分析。
的形态学分析。S1P的影响在收缩过程是评估通过计算pre-oligodendrocytes过程和成熟的少突胶质细胞的数量S1P-treated和控制文化。我们定义了三种类型的细胞,这取决于他们的过程:细胞没有过程,细胞有一个过程,与两个或两个以上的进程和细胞。结果表示为均值±SEM至少有三个独立的实验中,至少有三个文化/条件。使用学生的统计分析t测试。
免疫印迹分析。收获后,细胞与PBS冲洗,然后用1% SDS在50 m细胞溶解米三、pH值8.0和150米米包含1 m氯化钠缓冲区米衣饰归宿4和1米米PMSF。离心收集的溶解产物是15分钟在4°C。细胞溶解产物的蛋白质浓度是衡量BCA蛋白质测定(σ)。蛋白质(30μg)解决了10% sds - page电泳,electroblotted 1 h在4°C硝化纤维素膜(Amersham生物科学,Les乌里,法国)如前所述(Barbin et al ., 2004)。密度测量数据得到使用IQMac 1.2版本软件(分子动力学,桑尼维尔CA)。
RNA干扰。四个集中SMART-selected 21核苷酸RNA小干扰RNA (siRNA)工器,针对的核苷酸Edg8 / S1P5编码序列,设计并获得MWG生物技术(Courtaboeuf、法国)。小干扰rna进行瞬时转染的成熟少突细胞文化,3 d隔离后,使用jetSI (Qbiogen、Illkirch、法国),遵循制造商的指示。细胞治疗Edg8 / S1P5核(50 n米)前48 h S1P加法。控制文化nonrelevant对待MTLR-Cy3核(60 n米)(myotubularin加上Cy3) (MWG生物技术)。
结果
Edg8表达的类型/ S1P5产后大脑
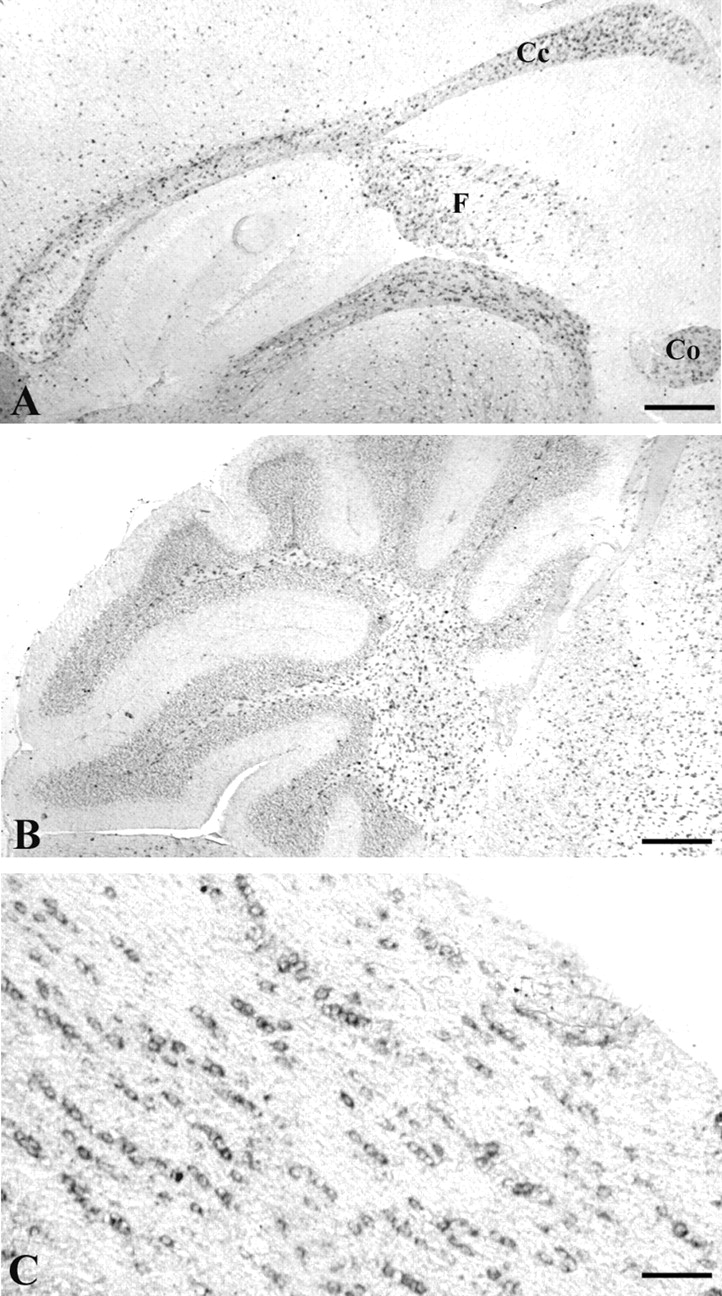
的表达Edg8 / S1P5分析了信使核糖核酸原位杂交的矢状部分老鼠大脑在一天产后21 (P21) (图2)。Edg8 / S1P5白质束表达细胞被发现,如胼胝体,伞,前连合,运行的白质束通过纹状体(图2一个)和小脑的深层白质(图2B)。白质束,Edg8 / S1P5mRNA-expressing细胞经常被观察到在一个连锁模式,高度提示intrafascicular寡树突胶质细胞(图2C)。

的表达Edg8 / S1P5信使核糖核酸体内。原位杂交在矢状digoxigenin-labeled P21的老鼠的大脑部分Edg8 / S1P5riboprobe (一个)表明,Edg8 / S1P5检测到mRNA在胼胝体(Cc),前连合(Co),伞(F)和小脑白质(B)。C白质束,Edg8 / S1P5mRNA-expressing细胞通常liein连锁模式,高度提示intrafascicular少突胶质细胞。规模的酒吧:A、B160μm;C80μm。
明确识别细胞的表型表达Edg8 / S1P5信使rna,我们利用1900 bp -MBP-lacZ转基因老鼠,大肠杆菌lacZ报告基因的控制下1.9 kb的初始部分MBP启动子(麻醉品et al ., 1992)。在这种转基因线,β-galactosidase记者表达的是成熟的髓鞘寡树突胶质细胞(Stankoff et al ., 1996)。矢状的部分P21老鼠大脑digoxigenin-labeled杂化Edg8 / S1P5riboprobe,其次是anti-β-galactosidase疣状标签少突细胞的细胞体(图3)。Edg8 / S1P5阳性细胞也anti-β-galactosidase正面的(图3A、B),因此确定为成熟的少突胶质细胞。

细胞定位Edg8 / S1P5-expressing细胞在成年老鼠的大脑。A、B矢状切面的MBP-lacZP21转基因小鼠大脑digoxigenin-labeled杂化Edg8 / S1P5riboprobe (一个与anti-β-galactosidase)和应用(B)。注意的表达Edg8 / S1P5(一个)β-galactosidase-positive(红色)成熟的少突胶质细胞(B)。C, D、双免疫染色Edg8 / S1P5 Ab (C)和anti-MBP马伯(D)显示colocalization Edg8 / S1P5和MBP免疫反应性。注意,与抗体,细胞的身体不检测。E, F、双免疫染色Edg8 / S1P5 Ab和anti-APC马伯,Edg8 / S1P5染色(绿色)似乎限制有髓纤维和APC未被检测到+少突细胞细胞体(红色)。规模的酒吧:A、B、E, F10μm;C, D500μm。
检查Edg8 / S1P5蛋白的表达在成熟的少突胶质细胞和髓鞘,成年人的大脑部分与anti-Edg8 double-labeled / S1P5 Ab和anti-MBP或anti-APC马伯。Edg8 / S1P5和MBP免疫反应性与检测和似乎仅限于有髓纤维(图3C, D)。然而,即使在强烈Edg8 / S1P5-positive有髓神经束,anti-Edg8 / S1P5 Ab未能标签APC-positive少突细胞细胞体(APC的标志成熟少突胶质细胞的胞体(Bhat et al ., 1996))(图3E, F)。这表明,类似于MBP, Edg8 / S1P5不积聚在髓鞘少突胶质细胞的胞体,而是转入流程和髓鞘。然而,Edg8 / S1P5沿着髓鞘(mRNA在不被察觉的情况下图3一个),这表明,形成鲜明对比mbp成绩单(安吉et al ., 1997),只有Edg8 / S1P5蛋白质,但不是它的信使rna,用这种方式运输。
进一步调查Edg8 / S1P5的表达,我们利用Edg8 / S1P5+ / -有一个等位基因的动物Edg8 / S1P5取而代之的是一个基因IRES-lacZ盒(图1)。因此,在这些杂合的小鼠,细胞表达记者β-galactosidase识别的细胞Edg8 / S1P5转录激活。双标记的成年Edg8 / S1P5+ / -老鼠大脑部分与anti-β-galactosidase抗血清和anti-Map-2 anti-GFAP或anti-MAC1和anti-F4 - 80抗体证明Edg8 / S1P5没有发现在神经元、星形胶质细胞或小胶质细胞,分别(数据没有显示)。此外,双immunolabeling anti-β-galactosidase和anti-NG2(信息公开化的标志(莱文和Stallcup, 1987;西山et al ., 1996)抗体的P5大脑部分Edg8 / S1P5+ / -老鼠显示NG2-positive细胞中,80%也β-galactosidase积极暗示的转录Edg8 / S1P5OPC阶段(图4)。观察,并非所有NG2-positive细胞β-galactosidase积极可能表明Edg8 / S1P5开始表达在OPC阶段。另外,因为喜欢的《忍者外传2》还提出了标签等大脑结构血管和小胶质细胞(妓女et al ., 1999;琼斯等人。,2002年),这些额外的NG2-positive /β-galactosidase-negative细胞可能代表不同于信息公开化。

Edg8 / S1P5 NG2-expressing表达的信息公开化。脑干矢状切面的P5产后Edg8 / S1P5+ / -哪一个鼠标Edg8 / S1P5等位基因是被一个取代IRES lacZ盒(见图1),显示。双重immunolabeling anti-β-galactosidase抗体(绿色;一个)和anti-NG2抗体(红色;B),合并C显示,Edg8 / S1P5mRNA-expressing细胞(沾anti-β-galactosidase抗体)NG2-positive公开化。比例尺,10μm。
表达Edg8 / S1P5少突细胞谱系的细胞文化
分析Edg8 / S1P5表达式进行浓缩oligodendroglial文化源自新生鼠脑,含有细胞oligodendroglial发展的不同阶段(Lubetzki et al ., 1991)。Edg8 / S1P5-expressing细胞被确认为A2B5+/ GalC- - - - - -信息公开化(图5A、B),O4+/ GalC- - - - - -preoligodendrocytes (图5C, D),或者分化GalC+不成熟的少突胶质细胞。此外,疣状的高纯度oligodendroglial文化源自成年鼠大脑(Lubetzki et al ., 1986)表明,PLP+成熟少突胶质细胞也Edg8 / S1P5+(图5E, F)。不像我们的发现在大脑的部分(图3),在这些文化中,Edg8 / S1P5蛋白质检测不仅在流程还在oligodendroglial细胞的胞体发育阶段检查。同意我们的观察大脑部分,Edg8 / S1P5特定oligodendroglial细胞免疫反应性,没有检测到SMI31/32染色+神经元,MAC1+/ f4 - 80+小胶质细胞,或GFAP-expressing星形胶质细胞(数据没有显示)。

Edg8 / S1P5表达少突细胞的文化。oligodendroglial文化源自新生鼠(模拟)和成年鼠大脑(E, F),Edg8 / S1P5蛋白(A, C, E)是在A2B5发现+信息公开化(B)和O4+pre-oligodendrocytes (D在PLP)以及+成熟的少突胶质细胞(F)。注意,Edg8 / S1P5细胞体和检测流程。比例尺,10μm
S1P促进过程收缩pre-oligodendrocytes通过激活Edg8 / S1P5
治疗S1P诱导pre-oligodendrocytes的生动形态变化。这种效应在文化源自新生鼠脑(Lubetzki et al ., 1991电镀后),3 d,高达65%的细胞O4-positive (图6一个)和GalC-negative pre-oligodendrocytes,平均四到六个流程的每一个细胞。添加S1P(1μ米30分钟诱导细胞四舍五入)(图6B),以90%的preoligodendrocytes收回所有的过程和剩余的细胞有一个进程左(7%)或两个或更多(3%)(见图8一个)。S1P过程收缩是剂量依赖性的影响,与突然转变S1P的浓度在0.01和0.1之间μ米(见图8一个)。S1P治疗引起的收缩的过程是短暂的,与流程reextension 48 h后更加明显在72 h (图6C, D)。Edg8 / S1P5激活的角色在这个收缩效应实验证实了文化源自Edg8 / S1P5- / -的大脑。在这些文化中,表型与野生型文化、S1P(1μ米)不产生收缩oligodendroglial流程(图6E, F)。这种效应的S1P oligodendroglial过程的收缩在成熟的少突胶质细胞(没有被观察到图7A、B),建议改变应对S1P pre-oligodendrocytes和成熟的少突胶质细胞之间。没有收缩并不是由于我们的文化条件,因为在控制实验中,我们证实,据Bagnard et al。(1998年3),添加semaphorin 24 h引发主要成熟的少突胶质细胞收缩的过程(图7C, D)。

pre-oligodendrocytes S1P诱发过程收缩。模拟,Preoligodendrocytes来自新生鼠脑治疗对S1P 30分钟(1μ米和O4 mAb)和应用。细胞形态学分析之前一个)和30分钟(B),48小时(C)和72 h (DS1P撤军后)。有一个戏剧性的损失过程发现添加S1P后30分钟,但过程reextension已经观察到48 h后。进程扩展72 h S1P加法后更明显。文化的少突细胞前体Edg8 / S1P5-deficient大脑前(E)和后30分钟(F)的S1P(1μ米)表明,在缺乏Edg8 / S1P5,没有过程收缩引起的S1P培养基。比例尺,10μm。

S1P-induced过程收缩不成熟的少突胶质细胞浓度依赖性,通过ρ激酶介导。一个,增加浓度的S1P被添加到未成熟少突细胞文化,有或没有ρ激酶抑制剂Y27632。细胞的数量过程分析了30分钟后。不同的列对应每个细胞的进程数量:0,□;1、▧;两个或两个以上的流程,▪。B,S1P诱导CRMP2在未成熟少突胶质细胞介导的磷酸化ρ激酶途径。不成熟的少突胶质细胞S1P处理(1μ米)15分钟,ρY27632激酶抑制剂的存在与否。Whole-celllysates与抗体制备和免疫印迹识别CRMP2的磷酸化形式。

缺乏S1P-induced成熟少突胶质细胞的收缩过程。成熟少突胶质细胞纯化从成年老鼠的大脑处理S1P(1μ米)(A、B)或3包含介质(某C, D)。少突胶质细胞的形态的影响,分析了应用anti-MBP抗体,之前A、CS1P)和24小时后(B3)或某(D)增加。3某,但不是S1P,诱发四舍五入成熟的少突胶质细胞。比例尺,10μm。
S1P-induced过程收缩preoligodendrocytes通过CRMP2磷酸化和ρ激酶介导
据报道,在神经细胞,生长锥崩溃引起的激活需要ρkinase-mediated LPA受体磷酸化CRMP2 (Arimura et al ., 2000)。解决的可能性CRMP2磷酸化参与S1P-induced oligodendroglial收缩过程,我们通过免疫印迹分析CRMP2磷酸化的提取物纯化O4 S1P-treated文化+/ GalC- - - - - -pre-oligodendrocytes或成熟PLP+少突胶质细胞,使用anti-phosphorylated CRMP2抗体。所示图8B提取物中检测出,磷酸化CRMP2文化pre-oligodendrocytes治疗15分钟S1P(1μ米)。相比之下,CRMP2磷酸化不是成熟的少突胶质细胞,引起S1P的协议没有S1P-induced过程中在这些细胞收缩。
确定S1P-induced CRMP2是由ρ激酶的磷酸化,我们检查了阻塞的影响ρ使用特定激酶激酶抑制剂Y27632ρ。添加Y27632前30分钟S1P除了导致CRMP2磷酸化的抑制(图8B),以及抑制S1P-induced过程的收缩(图8一个)。总之,这些结果表明,S1P-induced过程收缩pre-oligodendrocytes涉及CRMP2ρ激酶磷酸化。
S1P是成熟的少突胶质细胞的生存因素,通过Edg8 / S1P5
因为oligodendroglial文化的纯洁来自老鼠的大脑始终高于从老鼠的大脑,我们选择调查S1P的角色少突细胞在老鼠生存的文化。成熟少突胶质细胞分离成年大鼠大脑培养前3 d改变DM,缺乏胰岛素和FCS。文化DM条件下24小时后,我们观察到,肝癌和使用一个标准的可行性分析,减少了58%的成熟少突细胞生存与控制文化维持在一个中等补充胰岛素(10μ米)和FCS (0.5%)。相比之下,在相同的培养条件下,增加S1P DM导致显著增加少突细胞的生存。这种效应是浓度依赖性和1μ米浓度的S1P少突细胞的存活率几乎回到控制文化(图9一个)。S1P胰岛素和FCS-supplemented介质没有进一步增加成熟的少突胶质细胞的生存生存与控制[83和81% S1P(1μ米糖尿病和S1P)(1μ米分别)+胰岛素和FCS),表明没有添加剂的效果。效果观察与成熟的少突胶质细胞相比,没有变化的生存pre-oligodendrocytes S1P治疗O4后检测+富含文化(数据未显示)。

S1P诱发成熟少突胶质细胞的生存。成熟的少突胶质细胞,分离出还是老鼠大脑维持3 d在培养基含有胰岛素(10μ米)和FCS(0.5%),然后变成了DM(介质没有胰岛素和FCS)包含越来越S1P的浓度。细胞生存能力评估MTT生存分析,24 h后,结果表示为(即的比例控制。,细胞在胰岛素和FCS-supplemented介质)。去除胰岛素和FCS导致细胞生存能力(减少58%#p< 0.001 vs控制)。增加S1P DM导致浓度增加oligodendroglial生存(*p< 0.001 vs S1P未经处理的文化保持在DM)。添加1μ米从DM-induced S1P获救成熟少突胶质细胞受损的生存,生存回到水平在统计学上没有不同于控制细胞补充中。S1P的生存效果(1μ米)被添加PTX[°逆转p< 0.001 vs S1P(1μ米)对待文化没有PTX)。B在成熟的少突胶质细胞,S1P-induced一种蛋白激酶磷酸化。成熟少突胶质细胞被刺激1μ米20分钟的S1P PTX的存在与否。全细胞溶解产物与抗体制备和免疫印迹识别磷酸化Akt。C光密度数据均值±SEM的三个不同的实验。
确定生存效应引起的S1P被Edg8 / S1P5受体介导的信号,我们使用核执行功能丧失的实验。这个核过程,而不是从成人oligodendroglial文化Edg8 / S1P5- / -大脑,因为数量有限Edg8 / S1P5- / -老鼠。在初步实验中,使用nonrelevantMTLR-Cy3核,我们认为,在我们的实验条件下,转染效率的成熟(PLP)+)少突胶质细胞接近80% (图10一个),没有明显的细胞生存能力或形态学的变化。转染与Edg8 / S1P5siRNA导致水平的显著减少Edg8 / S1P5免疫反应性被西方墨点法。这种效果是非常具体的,因为没有减少的水平Edg2 /观察LPA1受体(图10B)。我们下一个检查添加S1P(1μ的效果米)的生存Edg8 / S1P5siRNA-treated成熟少突胶质细胞保持在DM。成熟的少突胶质细胞生存的增加引起的S1P文化nonrelevant对待MTLRsiRNA不再是中观察到Edg8 / S1P5siRNA-treated文化(图10C)。这些实验证明生存效应引起的S1P Edg8 / S1P5成熟的少突胶质细胞介导的受体。

Edg8 /小干扰rna沉默抑制S1P5 S1P-induced成熟少突胶质细胞的生存。一个,成熟的少突胶质细胞的转染MTLR-CY3核。少突胶质细胞是沾anti-GalC马伯(绿色)。箭头指向核,局部细胞体内(红点)。B,成熟的少突胶质细胞转染的浓度增加Edg8 / S1P5核。转染后48小时,全细胞溶解产物与抗体制备和免疫印迹识别Edg8 / S1P5或Edg2 / LPA1展示的具体表达抑制Edg8 / S1P5。C,成熟的少突胶质细胞转染Edg8 / S1P5核或nonrelevantMTLR核和维护在DM(见传说图9)。后48 h, S1P-mediated生存MTT生存分析,分析结果(即表示为的比例控制。S1P-treated细胞转染MTLRsiRNA DM和维护。#p< 0.001 vs控制)。
S1P-induced成熟少突胶质细胞的生存是通过Gα/我和Akt介导的通路
我们接下来研究的信号转导机制激活Edg8 / S1P5 S1P受体介导的成熟的少突胶质细胞。首先,我们表明,S1P-induced生存抑制了先前与PTX治疗的文化,认为这一现象是介导通过PTX-sensitive Gα/ i-protein (图9一个)。因为前面所显示的一种蛋白激酶信号级联一直参与少突细胞生存(Vemuri McMorris, 1996;弗洛雷斯et al ., 2000),我们执行一个免疫印迹anti-phosphoAkt马伯S1P-treated细胞提取物成熟少突胶质细胞。暴露15分钟的成熟的少突胶质细胞S1P诱导一个健壮的一种蛋白激酶的磷酸化,这主要是通过与PTX孵化的细胞(图9B, C)。相比之下,文化丰富的S1P治疗O4+细胞未能产生一种蛋白激酶磷酸化pre-oligodendrocytes(数据没有显示)。
讨论
Oligodendroglial Edg8 / S1P5的表情
在他们最初的报告识别Edg8 / S1P5 Im et al。(2001年)表示,在大脑中,Edg8 / S1P5主要观察mRNA在白质束,暗示表达在成熟的少突胶质细胞,证实了在文化分化少突胶质细胞(Yu et al ., 2004)。这是与发现的毡帽et al。2003年)报道Edg8 / S1P5 NG2-positive表达的蛋白质信息公开化但不成熟的少突胶质细胞。在我们的实验条件下,我们显示,两种都用原位杂交和immunolabeling Edg8 / S1P5的表达在成熟的少突胶质细胞的老鼠的大脑。同意报告的毡帽et al。2003年),我们也能够发现Edg8 / pre-oligodendrocytes S1P5表情和大部分的信息公开化。这些数据符合整个oligodendroglial Edg8 / S1P5发展的一个表达式。
激活Edg8 pre-oligodendrocytes / S1P5诱发过程收缩
我们表明,S1P oligodendroglial前兆的刺激,但不成熟的少突胶质细胞,导致了oligodendroglial过程收缩Edg8 / S1P5受体介导的。在类似的方式,LPA引起收缩过程的不成熟,但不成熟,CG4 oligodendroglial细胞系和信息公开化道森et al ., 2003),效果很可能由Edg2 / LPA1受体(Allard et al .,1998年,1999年)。的注意的是,在相同的研究中,道森et al。(2003年)报道,除了LPA, S1P也诱导未分化CG4细胞的收缩过程。
S1P的行为和以初级神经LPA文化已经被许多组织和显示特征诱导神经突生长锥收缩和/或崩溃(佐藤et al ., 1997;Hirose et al ., 1998;福岛et al ., 2002)。后续的研究涉及的参与一个丝氨酸/ threonine-dependentρ激酶在lysophospholipid-induced神经收缩和识别肌球蛋白轻链和CRMP2为ρkinase-mediated主要基质收缩。这种转导级联因此被建议作为一个关键系统lysophospholipid-induced生长锥崩溃和被认为是轴突生长锥的一个重要途径改造(Moolenaar 1995;梁et al ., 1996;Ishizaki et al ., 1997;Arimura et al ., 2000)。我们的研究结果表明,同一通路与S1P-induced preoligodendrocyte过程的收缩,这效果是依赖于ρ激酶的活性和增加CRMP2的磷酸化。Semaphorin 3 a,一个已知的chemorepellent公开化迁移到视神经(杉本et al ., 2001;斯帕斯基et al ., 2002),也称诱导过程收缩少突胶质细胞。然而,改装车3 a-induced oligodendroglial流程的收缩,通过CRMP2介导,发现被限制在成熟的少突胶质细胞(里卡德et al ., 2000)。这些结果表明,S1P和某3 pre-oligodendrocytes可以诱导oligodendroglial过程收缩和成熟的少突胶质细胞,分别。这些配体施加他们的行动与不同受体交互和Edg8 / 3 (1 neuropilin Sema S1P5 S1P)表示pre-oligodendrocytes和成熟的少突胶质细胞,和CRMP2下游信号通路的一个常见的元素。
的功能后果S1P-mediated过程收缩pre-oligodendrocytes仍然投机。可能在迁移之后pre-oligodendrocytes解决在给定在启动前束轴突髓鞘形成的过程。髓鞘形成过程(即。,the enwrapping of each axon segment by a single oligodendrocyte extension) is preceded by a major modification of the morphology of oligodendrocytes. Both在活的有机体内和在体外,它已经表明,这个终端分化少突细胞的特点是数量急剧减少的过程,把星状,非常明亮的,premyelinating少突细胞变成一个成熟髓少突细胞糖蛋白(MOG)表达髓鞘与几个主要过程少突细胞,髓鞘将年底延长(Lubetzki et al ., 1993;普费弗et al ., 1993;萨利et al ., 1996)。在这方面,一个潜在的功能Edg8 / S1P5受体少突胶质细胞可以调节分枝减少,这样的数量少突细胞过程匹配需要髓鞘形成的轴突段的数量。也感兴趣的似乎有一定程度的冗余系统中,S1P和LPA导致信息公开化的收缩过程。这一发现或许可以解释缺乏明显的髓磷脂病理学在转基因线缺陷Edg2 / LPA1(来自et al ., 2002)或Edg8 / S1P5(补充图S1,可用www.jneurosci.org作为补充材料)。在这方面,它将感兴趣的检查的表型Edg2 / LPA1- / -/ Edg8 S1P5- / -小鼠的两倍。
成熟的少突胶质细胞S1P和生存
如前所述,S1P成熟少突胶质细胞的激活对收缩过程没有影响。然而,我们观察到显著和显著增加成熟少突细胞生存(即PTX敏感。Gα/我激活)并通过Akt介导的信号通路。在这方面,我们的结果与先前协议确定角色Akt激活和生存的S1P肝myofibroblast或内皮细胞(Morales-Ruiz et al ., 2001;Davaille et al ., 2002)。各种不同的代理已被证明会促使少突细胞生存,和一些如NT3 IGF-I和调节(Vemuri McMorris, 1996;弗洛雷斯et al ., 2000通过磷脂酰肌醇3′)信号激酶(PI3K)转导通路。S1P因此似乎是另一个oligodendroglial生存因素,通过一种蛋白激酶信号通路下游Edg8 / S1P5受体。
我们的研究结果显示,在oligodendroglial谱系的细胞,激活Edg8 / S1P5 S1P受体可以触发两种截然不同的生理效应,取决于细胞的发育阶段。在pre-oligodendrocytes S1P收缩的过程是由一种ρ激酶/ CRMP2通路,而在成熟的少突胶质细胞,通过PTX-sensitive S1P促进生存途径。很可能这些通路被激活不同的g,和Edg8 / S1P5已被证明夫妻Gαi和G12 (马列et al ., 2001)。PI3K通路已被证明是由Gαi和ρ激酶途径激活需要G12/13 (李et al ., 2003)。功能变化的影响Edg8 / S1P5激活可能因此与受体的变化耦合与heterotrimeric g。这可能发生由多种蛋白表达的潜在机制包括发育开关(即。,from G12/13 to Gα expression) or alternative splicing of the Edg8/S1P5 receptor resulting in changes to the coupling efficiency of the receptor to specific G-proteins, as suggested for Edg2/LPA1 (Allard et al ., 1999)。然而,我们无法孤立另一种拼接的变体Edg8 / S1P5相比之下的RNA提取O4+纯度的浓缩铀pre-oligodendrocytes和成熟的少突胶质细胞(未发表的数据)。我们的研究结果是一致的与观测Edg2 / LPA1-expressing雪旺细胞,其中Edg2 / LPA1激活LPA介导许旺细胞生存通过PI3K /增殖kinase-dependent通路和细胞骨架变化引发的ρkinase-dependent通路(维纳和春,1999;李et al ., 2003)。
在多发性硬化症,remyelination退化引起轴突疾病过程往往失败。Oligodendroglial灭绝可能占这个修复赤字比例的病变特点是少突细胞损失。然而,没有髓鞘修复也被报道与幸存的少突细胞祖细胞病变或分化少突胶质细胞,这表明这些细胞无法或被阻止形成髓鞘(Chang et al ., 2002;富兰克林,2002;雷诺兹et al ., 2002)。因此,理解调制的信号通路的激活和失活髓鞘过程可能使小说蛋白质的识别,最终可能允许的调制改善这种毁灭性疾病的疗法。这里提供的数据表明,Edg8 / S1P5受体可能就是这样一个蛋白质,参与髓鞘过程的监管。通过中介少突胶质细胞的发育调节dual-signaling通路,导致两个截然不同的功能影响调制过程收缩或细胞生存,Edg8 / S1P5受体的激活可能是重要的少突胶质细胞的成熟和生存和最终可能发挥关键作用的调制,髓鞘的形成,和修复。
脚注
这项工作是由国家卫生研究所et de la医学研究院和协会de矫揉造作的苏尔la斑块硬化。C.J.的接受者是葛兰素史克(GSK)奖学金。我们感谢Drs。v·柯林斯k . Ikenaka b . Ranscht b .骗子和j . Trotter的礼物价值的试剂,安娜·威廉姆斯博士仔细阅读手稿,和葛兰素史克公司实验动物科学和发现管道遗传学团队畜牧业和基因分型。
信件应该向凯瑟琳Lubetzki博士生物神经元/ Glie des交互,国家卫生研究所et de la医学研究院联合Mixte de矫揉造作的- 711,友谊医院de la弗尔f - 75651巴黎,法国。电子邮件:catherine.lubetzki在{}psl.ap-hop-paris.fr。
版权©2005年神经科学学会0270 - 6474/05/251459 - 11美元的15.00/0
↵*C.J.和萨达姆政权造成了同样的工作。








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}