

文摘gydF4y2Ba
研究表明,有有益的锻炼在帕金森症患者,但潜在的分子机制负责这些影响知之甚少。在啮齿动物模型研究提供了一种方式来检查运动对多巴胺能神经传递的影响。使用密集的跑步机锻炼,我们决定改变1-methyl-4-phenyl-1纹状体多巴胺,2,3,6-tetrahydropyridine注射(MPTP药物)进行鼠标。C57BL / 6 j小鼠被分为四组:(1)盐,(2)生理盐水+运动,注射注射(3)MPTP药物,和(4)MPTP药物+运动。运动开始注射后5 d MPTP药物损伤和持续了28 d。电动机速度的跑步机上跑步均得到提升运动组。行使所有动物也显示增加延迟下降(改善)使用加速rotarod处理后与未处理的老鼠。使用高效液相色谱法,我们没有发现注射在纹状体多巴胺组织水平差异MPTP药物+运动小鼠注射了MPTP药物相比。有一个增加盐水+运动小鼠中发现。使用快速扫描环绕伏安测量法分析显示增加stimulus-evoked释放和减少注射衰变的背侧纹状体的多巴胺MPTP药物+运动只老鼠。免疫组织化学染色分析纹状体酪氨酸羟化酶和多巴胺转运蛋白显示减少注射表达MPTP药物+运动小鼠注射了MPTP药物老鼠相比。 There were no differences in mRNA transcript expression in midbrain dopaminergic neurons between these two groups. However, there was diminished transcript expression in saline plus exercise compared with saline mice. Our findings suggest that the benefits of treadmill exercise on motor performance may be accompanied by changes in dopaminergic neurotransmission that are different in the injured (MPTP-lesioned) compared with the noninjured (saline) nigrostriatal system.
介绍gydF4y2Ba
从几个实验室包括我们的最近的研究表明,运动可以产生有益的影响帕金森病(PD)患者和PD在啮齿动物模型(gydF4y2Ba考米拉et al ., 1994gydF4y2Ba;gydF4y2Ba蒂勒森et al ., 2001gydF4y2Ba,gydF4y2Ba2003年gydF4y2Ba;gydF4y2BaBezard et al ., 2003gydF4y2Ba;gydF4y2Ba费舍尔et al ., 2004gydF4y2Ba;gydF4y2Ba陈et al ., 2005gydF4y2Ba;gydF4y2BaFaherty et al ., 2005gydF4y2Ba)。在6-hydroxydopamine (6-OHDA)损伤鼠和1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine注射(MPTP药物)进行鼠标,运动开始之前或期间neurotoxicant接触已被证明是神经保护,纹状体多巴胺损失的衰减做了演示。我们之前的研究表明,高强度的跑步机锻炼,发起5 d后注射管理MPTP药物,时期neurotoxicant-induced完成细胞死亡,也会导致改善电机性能注射的MPTP药物损伤小鼠(gydF4y2BaJackson-Lewis et al ., 1995gydF4y2Ba;gydF4y2Ba费舍尔et al ., 2004gydF4y2Ba)。此外,多巴胺转运体(DAT)的差别,我们发现对这些基因,蛋白质重要调节多巴胺的吸收,和upregulation多巴胺受体D2受体的激活是重要的诱发运动行为。鉴于多巴胺的重要作用在运动学习和执行,和我们兴趣受伤的基底神经节的神经可塑性,在这项研究中,我们审查的影响运动对纹状体多巴胺能系统的其他潜在的补偿变化包括总纹状体多巴胺水平和释放。我们进一步检查是否运动给蛋白质重要改变多巴胺生物合成(酪氨酸羟化酶(TH)、吸收(DAT)可能是伴随着各自的mRNA转录表达的改变在中脑多巴胺能神经元和运动是否会导致黑质神经元的数量的变化。gydF4y2Ba
这些研究中,我们使用四组老鼠包括(1)盐,(2)生理盐水+运动,注射注射(3)MPTP药物,和(4)MPTP药物+运动。密集的跑步机上跑步开始注射后5 d MPTP药物损伤和持续了28 d运行(5 d /周)。从所有四组脑组织在完成练习。分析包括(1)纹状体多巴胺及其代谢产物的测量单位,(2)纹状体多巴胺释放在片文化中使用快速扫描环绕伏安测量法。黑多巴胺能神经元的无偏stereological计数被用来确定组之间的任何差异在细胞数量。我们的研究显示,锻炼可能有一个微分影响多巴胺能系统MPTP-versus saline-treated老鼠。的mptp缺陷型小鼠中,运动对多巴胺的释放产生影响,这是特定的区域,但不是总纹状体多巴胺水平。saline-treated老鼠,运动增加总纹状体多巴胺对多巴胺的释放水平没有显著影响。此外,在我们的研究中,我们注射确定MPTP药物管理导致细胞损失50%,注射在黑质细胞数量之间没有区别MPTP药物注射和MPTP药物+运动的老鼠。Downregulation DAT和TH转录表达的只是在生理盐水+ exercise-treated发现老鼠。gydF4y2Ba
材料和方法gydF4y2Ba
动物。gydF4y2Ba
老鼠用于这些研究是年轻人(8 - 10周大)雄性C57BL / 6 j小鼠提供从杰克逊实验室(巴尔港,我)。有四个治疗组包括以下几点:(1)盐,(2)生理盐水+运动,注射注射(3)MPTP药物,和(4)MPTP药物+运动。动物被安置五到笼子里,适应12 h光/暗周期的转变,这样运动发生在动物正常后时期。所有实验进行按照美国国立卫生研究院(NIH)gydF4y2Ba指导实验室动物保健和使用的gydF4y2Ba(NIH出版80 - 23,1996年修订)和南加州大学的批准机构动物保健和使用委员会。gydF4y2Ba
MPTP损伤。gydF4y2Ba
MPTP(σ,圣路易斯,密苏里州)管理的一系列四腹腔内注射20毫克/公斤(自由基地)每隔2 h共管理80毫克/公斤。这种方案会导致∼60%黑神经元的损失,由无偏stereological TH染色和尼氏小物质和技术的80 - 90%损耗纹状体多巴胺水平(gydF4y2BaJackson-Lewis et al ., 1995gydF4y2Ba;gydF4y2BaJakowec et al ., 2004gydF4y2Ba)。黑细胞注射完成后,第三天MPTP药物损失政府决定通过计算剩余的黑TH免疫反应性的退化的神经元细胞,减少银染色(gydF4y2BaJackson-Lewis et al ., 1995gydF4y2Ba;gydF4y2BaJakowec et al ., 2004gydF4y2Ba)。gydF4y2Ba
跑步机锻炼。gydF4y2Ba
开始的前一周跑步机运动范式,六十老鼠可以保持前沿45厘米为5分钟跑步机带5.0米/分钟被随机分配到四组,以确保所有的动物在跑步机上执行同样任务注射前MPTP药物损伤。在这些研究中使用的跑步机是一个模型EXER-6M跑步机由哥伦布乐器制造(哥伦布,哦)。non-noxious刺激(metal-beaded窗帘)被用作触觉刺激,防止动物在跑步机上。因此,shock-plate激励是不习惯和压力相关的活动被最小化。运动发起5 d注射生理盐水或MPTP药物损伤后细胞死亡是完整的。老鼠从每个两个运动组(生理盐水+注射运动和MPTP药物+运动)是同时运行的六车道跑步机。运动持续时间30分钟开始逐步增加1天到达目标两个交易日持续时间30分钟(60分钟),每5 d /周(5分钟预热期),共有28 d运动(对应于最后一次注射后的42 d MPTP药物损伤)。跑步机的速度和运动时间为每个组增加时每组内的所有老鼠保持前沿在跑步机上皮带运行期间的75%。控制任何跑步机跑步做运动的影响(处理、小说环境噪声和振动),做运动组放置在顶部的跑步机时间相当于运动训练器(gydF4y2BaFukai et al ., 2000gydF4y2Ba;gydF4y2BaKojda et al ., 2001gydF4y2Ba)。gydF4y2Ba
电动机与rotarod行为分析。gydF4y2Ba
加速rotarod(五车道rotarod加速;尤格Basile Comerio、意大利)是用来测量电机平衡和协调在跑步机上运动期间。rotarod由一个旋转主轴(直径3厘米),老鼠挑战速度和交替旋转方向。杆的速度设置为30 rpm(正反向)每24 s的变化方向。每个老鼠被放置在一个单独的隔间旋转杆和延迟下降是由磁自动记录旅行盘子。所有组的老鼠每周测试一次。开始后的第一个星期记录3 d跑步机锻炼。连续5试验的测试由相隔1分钟,最高截止200年代的延迟。每个老鼠每周数据随后被收集,直到完成运动范式(对应于周6)。gydF4y2Ba
组织收集。gydF4y2Ba
脑组织从所有组老鼠收集最后一天的运动(28天的运动相应的注射后42 d MPTP药物损伤)。纹状体脑组织也收集了从每个实验组动物的一个子集的5 d运动(10 d postlesioning)来确定程度的注射多巴胺损耗在早期运动和MPTP药物时间点。老鼠被颈椎错位对新鲜组织(快速扫描环绕伏安测量法,高效液相色谱法gydF4y2Ba原位gydF4y2Ba杂交组织化学)或戊巴比妥紧随其后transcardial灌注与固定剂(免疫组织化学和无偏stereological计数)。纹状体组织进行高效液相色谱分析收集新鲜全体对应于解剖区域从1.20前囱前囱0.60,与边界背前连合,腹侧胼胝体,内侧侧脑室,和2.5毫米侧中线和冷冻,直到分析。gydF4y2Ba原位gydF4y2Ba杂交组织化学、快速扫描环绕伏安法和免疫组织化学进行冠状部分对应于1.30前囱前囱0.00。此外,最初评价程度的MPTP-mediated纹状体多巴胺耗竭,脑组织用于高效液相色谱分析收集10 d post-MPTP损伤的一个子集做老鼠注射生理盐水和MPTP药物组。gydF4y2Ba
高效液相色谱法分析多巴胺及其代谢产物。gydF4y2Ba
根据改编自神经递质浓度测定gydF4y2Ba欧文et al。(1992)gydF4y2Ba的方法gydF4y2Ba基尔帕特里克et al。(1986)gydF4y2Ba。组织分析均质在12000×0.4 N高氯酸和离心机gydF4y2BaggydF4y2Ba分离沉淀蛋白质。蛋白质颗粒在0.5 N resuspended氢氧化钠和总蛋白浓度决定使用Coomassie +蛋白质分析系统(皮尔斯,罗克福德,IL)使用Biotek模型Elx800标(Wincoski Biotek乐器,VT)和KCjunior软件。多巴胺的浓度,3,4-dihydroxyphenylacetic (DOPAC)和高香草酸(HVA)化验通过高效液相色谱电化学检测。样本注入一个ESA autosampler(切姆斯福德,MA)。多巴胺及其代谢产物分离的150×3.2 mm反向阶段3-μm-diameter C-18列(ESA)监管在28°C。流动相MD-TM (ESA)由acetylnitrile磷酸盐缓冲剂和离子对代理交付0.6毫升/分钟的速度。电化学检测器是ESA模式Coularray 5600一套四通道分析细胞和三个潜力在100−,50和220 mV。高效液相色谱法是与Dell gx - 280计算机集成与分析项目包括ESA Coularray Windows软件和统计包InStat (GraphPad软件,圣地亚哥,CA)。gydF4y2Ba
快速扫描环绕伏安测量法测量多巴胺释放的大脑切片。gydF4y2Ba
快速扫描环绕伏安测量法用于分析多巴胺释放冠状,gydF4y2Ba在体外gydF4y2Ba纹状体脑片(gydF4y2Ba帕特尔和大米,2006年gydF4y2Ba)。大脑被放置在冷却(1 - 4°C),修改,和含氧人工脑脊液(aCSF)包含以下(mgydF4y2Ba米gydF4y2Ba):124氯化钠1.3 MgSOgydF4y2Ba4gydF4y2Ba1.25,3.0氯化钾,不gydF4y2Ba2gydF4y2Ba阿宝gydF4y2Ba4gydF4y2Ba26 NaHCOgydF4y2Ba3gydF4y2Ba2.4 CaClgydF4y2Ba2gydF4y2Ba10.0葡萄糖平衡95% OgydF4y2Ba2gydF4y2Ba/ 5%股份有限公司gydF4y2Ba2gydF4y2Ba混合获得的pH值7.3 - -7.4。修改aCSF,一些钠取代蔗糖来减少组织兴奋性在大脑切片切(蔗糖124米gydF4y2Ba米gydF4y2Ba,氯化钠62gydF4y2Ba米gydF4y2Ba)来维持正常的渗透压平衡aCSF。Hemicoronal纹状体片被削减的喙的组织厚度400年底μm Vibratome 1000 (Vibratome、圣路易斯、钼)。片被立即放置在含氧aCSF慢慢室温(23°C)。片仍在解决方案2 h和录音。单片都被转移到记录室(哈斯坡道风格气体界面室)与含氧aCSF沐浴不断解决方案的温度保持在32°C。盘碳纤维微电极(cfm)是由7毫米无浆碳纤维(德文郡,格拉汉姆·古德费勒公司PA)电泳阳极沉积涂料(gydF4y2Ba舒尔特Chow, 1996gydF4y2Ba)。细胞外多巴胺在发动机工厂监控每100毫秒运用三角波形(−0.4 + 1.0 V vs Ag / AgCl, 300 V / s)。电流记录与修改VA-10×伏安和电流放大器(NPI电子,塔姆,德国)。数据采集是由Clampex 7.0软件(分子装置,门洛帕克,CA)。电刺激是用来引起多巴胺与扭曲的流出,双相,镍铬合金电极片的表面。单脉冲(0.1毫秒,200μA)生成与Master-8脉冲发生器(AMPI,耶路撒冷,以色列)。200年恒流μA和0.1毫秒时间是通过使用一个A360R恒流刺激隔离器(世界精密仪器,萨拉索塔,FL)。刺激脉冲之间的间隔不是< 5分钟。cfm是75 - 100μm插入位置的切片100 - 200μm刺激电极对(gydF4y2Ba英里et al ., 2002gydF4y2Ba)。多巴胺在五个地点取样,每个薄片代表内侧侧和背腹侧维度。三个吻侧片检查在每个鼠标和每个动物的平均价值。改变细胞外多巴胺测定通过监测当前在200 mV窗口峰值氧化潜力多巴胺。Background-subtracted循环voltammograms是由减去当前获得之前从当前获得的刺激多巴胺的存在。将氧化电流转化为多巴胺浓度,电极与多巴胺标准校准实验使用后的解决方案。诱发多巴胺释放的机制在我们的伏安法实验测试一组大脑切片通过刺激纹状体组织使用以下序列变化的解决方案:控制aCSF包含2.4米gydF4y2Ba米gydF4y2BaCagydF4y2Ba2 +gydF4y2Ba和1.3gydF4y2Ba米gydF4y2Ba毫克gydF4y2Ba2 +gydF4y2Ba其次是减少CagydF4y2Ba2 +gydF4y2BaaCSF解决方案包含0.5米gydF4y2Ba米gydF4y2BaCagydF4y2Ba2 +gydF4y2Ba和3.2gydF4y2Ba米gydF4y2Ba毫克gydF4y2Ba2 +gydF4y2Ba与控制,其次是冲刷aCSF解决方案,然后以控制aCSF解决方案包含1μgydF4y2Ba米gydF4y2Batetrototoxin (TTX)。gydF4y2Ba
由intrastriatal刺激所引起的多巴胺信号的动力学研究了监测循环伏安法信号1 s之前和5 s intrastriatal刺激后的采样率每隔100毫秒(10 Hz)。多巴胺信号的衰减是由正常化postpeak多巴胺测量峰值测量多巴胺。衰变常数是决定从单一指数衰变的多巴胺信号根据以下方程:gydF4y2BaygydF4y2Ba=gydF4y2BaAegydF4y2Ba−ktgydF4y2Ba,在那里gydF4y2Ba一个gydF4y2Ba的多巴胺信号峰值时间0和常数−gydF4y2BakgydF4y2Ba是指数衰减的衰变率的多巴胺信号。方差分析进行所有组之间的衰变速率常数(−gydF4y2BakgydF4y2Ba)(gydF4y2BaMosharov苏尔寿公司,2005年gydF4y2Ba)。gydF4y2Ba
免疫组织化学染色。gydF4y2Ba
纹状体的相对表达TH免疫反应性(TH-ir)和DAT免疫反应性测定使用商用主要抗体在组织的部分,包括兔多克隆anti-TH和鼠标单克隆anti-DAT(微孔,贝德福德,MA)使用验证与比较相对的表达式所描述的模式gydF4y2Ba伯克et al。(1990)gydF4y2Ba。主要抗体绑定是可视化使用荧光标记二次抗体AlexaFluor 680或800 AlexaFluor (Licor、林肯、NE)。以确保染色强度的差异是由于抗原表达的差异,从每个不同的多个部分治疗组在相同的染色条件下同时处理。控制实验排除主要抗体或二次抗体也执行验证染色特异性。图像分析,三个或四个动物每治疗组和10 - 12部分动物被捕低倍镜下和数字化。相对光密度(表示为任意单位检测的线性范围内)的背侧纹状体是由减去胼胝体的相对光密度作为背景。gydF4y2Ba
无偏stereological计数的多巴胺能神经元。gydF4y2Ba
黑的数量在黑质致密部多巴胺能神经元(SNpc)确定使用无偏体视学和计算机成像程序BioQuant新星' (BioQuant成像,纳什维尔,TN)和一个奥林巴斯BX-50显微镜(奥林巴斯光学、东京、日本)配备一个机动阶段和数字Retiga-cooled CCD相机(Q-Imaging,本拿比,不列颠哥伦比亚,加拿大)。从三个每组小鼠脑组织制备。组织切片在30μm厚度和每个第六部分收集和彩色TH-ir使用兔多克隆抗体(微孔)和尼氏小物质复染色(gydF4y2BaJakowec et al ., 2004gydF4y2Ba;gydF4y2BaPetzinger et al ., 2005gydF4y2Ba)。SNpc是描述基于TH-ir从其余的大脑。部分集开始吻侧黑质在前囱−2.50毫米关闭前的第三脑室到桥的原子核的突出前囱−4.24毫米根据立体定位小鼠大脑的阿特拉斯(gydF4y2Ba2001年Paxinos和富兰克林gydF4y2Ba)。每个彩色腹侧中脑部分被低倍镜下(10×目标)和SNpc概述划定使用第三个从腹侧tegmental-immunoreactive神经元神经和大脑脚作为地标。神经元被认为高倍镜(80×目标)和计算如果他们显示TH-ir,都有一个明确的核,细胞质和细胞核。SNpc多巴胺能神经元的数量确定方法的基础上gydF4y2Ba甘德森和詹森(1987)gydF4y2Ba。gydF4y2Ba
原位gydF4y2Ba杂交组织化学。gydF4y2Ba
大脑的gydF4y2Ba原位gydF4y2Ba杂交很快被删除并冻结在异戊烷在干冰,和组织处理如前所述(gydF4y2BaJakowec et al ., 1995gydF4y2Ba,gydF4y2Ba2004年gydF4y2Ba)。选定的幻灯片是蘸NTB-2(柯达,罗彻斯特,纽约)照相乳剂,D-19开发的开发人员,和反沾甲苯基紫罗兰。最小化潜在的差异来源不同的实验,比较幻灯片都加工在同一实验中使用相同的杂交缓冲,探针浓度、调查准备、清洗方案,和乳液。中脑细胞捕获的图像使用奥林巴斯BX-51显微镜和计算机图像分析程序BioQuant (BioQuant成像)。乳胶粒数高于单个神经元内SNpc如果他们显示彩色细胞质和细胞核明显。gydF4y2Ba
统计分析的数据。gydF4y2Ba
统计分析了使用SPSS 14.0版Windows (SPSS,芝加哥,IL)或InStat软件(GraphPad软件)。行为测试组之间的差异进行了分析使用重复测量方差分析与主题之间的因素被病变(注射生理盐水或MPTP药物)和干预(运动或不运动)和在主体因素是时间。高效液相色谱分析、免疫细胞化学染色和颗粒计数gydF4y2Ba原位gydF4y2Ba杂交组织化学,双向方差分析比较不同的组织和检查执行重要的交互。gydF4y2Ba事后gydF4y2Ba与Bonferroni调整的轨迹进行,以确定任何显著差异。多巴胺释放的研究使用的数据快速扫描环绕伏安测量法进行了分析使用重复测量方差分析与主题之间的因素被病变(注射生理盐水或MPTP药物)和干预(运动或不运动)和被试因素电极位置(位置1 - 5)。gydF4y2Ba事后gydF4y2Ba在这些情况下,保证执行测试。试使用Huynh-Feldt校正效果进行球形。对所有分析,显著性水平gydF4y2BapgydF4y2Ba使用< 0.05。gydF4y2Ba
结果gydF4y2Ba
改善的时间进程的运行速度注射生理盐水+运动和MPTP药物+运动团体的28 d跑步机跑步所示gydF4y2Ba图1gydF4y2Ba。生理盐水+运动小鼠在第一天开始注射13.3米/分钟和MPTP药物+运动小鼠在第一天开始在7.6米/分钟。在跑步机上跑步的第一个星期,生理盐水组速度增加到14±1.4 m / min,进一步增加到22.6±0.3米/分钟的最后一周。注射的MPTP药物+运动组的运行速度增加到9.2±1.1米/分钟在第一周,进一步增加到20.5±0.7米/分钟在上周。如我们之前的研究中,所示gydF4y2Ba事后gydF4y2Ba分析显示出显著差异速度之间的第一天注射生理盐水+运动和MPTP药物+运动团体和这种差异不显著的完成跑步机上运行方案(gydF4y2Ba费舍尔et al ., 2004gydF4y2Ba)。小鼠注射的运行速度MPTP药物做最后一天的跑步机运动类似于注射的速度MPTP药物+运动组第一天的运动方案(数据未显示)gydF4y2Ba费舍尔et al ., 2004gydF4y2Ba)。gydF4y2Ba

电动跑步机上运动行为的分析。生理盐水和的mptp缺陷型小鼠都运行在电动跑步机上28 d(每周5 d)。的运行速度在每组小鼠(gydF4y2BangydF4y2Ba= 12)测定三次每周和比较。图表表明,(1)的mptp缺陷型小鼠有运行速度小于生理盐水组,星号显示显著差异(gydF4y2BapgydF4y2Ba< 0.05),(2)盐水和mptp缺陷型组织提高运行速度,最后两周,两组之间的差异不显著(gydF4y2Ba事后gydF4y2Ba分析;gydF4y2BapgydF4y2Ba< 0.05;gydF4y2BatgydF4y2Ba测试)。误差线表明SEM。gydF4y2Ba
作为第二测量电动机的性能,从所有四组老鼠进行测试的延迟从加速rotarod下降(gydF4y2Ba图2gydF4y2Ba)。因为高度的挑战这个任务,老鼠从所有组最初表现不佳。然而,我们观察到加班,有显著影响的运动(跑步)意味着延迟从加速rotarod(下降(以秒为单位)gydF4y2BaFgydF4y2Ba(44)gydF4y2Ba= 9.587;gydF4y2BapgydF4y2Ba< 0.0001)。注射两MPTP药物+运动和生理盐水+运动小鼠的表现要好于rotarod处理后比未处理组。没有显著影响注射的MPTP药物意味着延迟下降(gydF4y2BaFgydF4y2Ba(44)gydF4y2Ba= 0.851;gydF4y2BapgydF4y2Ba= 0.504)和注射之间没有显著的交互运动和MPTP药物意味着延迟下降(gydF4y2BaFgydF4y2Ba(44)gydF4y2Ba= 0.965;gydF4y2BapgydF4y2Ba= 0.435)。gydF4y2Ba

分析行为rotarod老鼠从所有组每周测试一次延迟下降的加速rotarod(均值和SEM每周以秒为单位)。有显著影响的运动(跑步)的平均延迟下降加速rotarod(以秒为单位),其中星号表示重要的运动,没有运动组之间要与众不同(gydF4y2BaFgydF4y2Ba(44)gydF4y2Ba= 9.587;gydF4y2BapgydF4y2Ba< 0.0001)。注射两MPTP药物+运动和生理盐水+运动小鼠的表现要好于rotarod处理后比未处理组。没有显著影响注射的MPTP药物意味着延迟下降(gydF4y2BaFgydF4y2Ba(44)gydF4y2Ba= 0.851;gydF4y2BapgydF4y2Ba= 0.504)和注射之间没有显著的交互运动和MPTP药物意味着延迟下降(gydF4y2BaFgydF4y2Ba(44)gydF4y2Ba= 0.965;gydF4y2BapgydF4y2Ba= 0.435)。gydF4y2Ba
高效液相色谱法分析是用来确定纹状体多巴胺的水平,其代谢物HVA和DOPAC,代谢物周转率,定义如下:[(DOPAC + HVA) /多巴胺)。这些数据所示gydF4y2Ba表1gydF4y2Ba和gydF4y2Ba图3gydF4y2Ba。首先,锻炼5 d组,有显著的影响注射的MPTP药物多巴胺的水平(gydF4y2BaFgydF4y2Ba(16)gydF4y2Ba= 39.52;gydF4y2BapgydF4y2Ba(< 0.0001)和多巴胺营业额gydF4y2BaFgydF4y2Ba(16)gydF4y2Ba= 88.30;gydF4y2BapgydF4y2Ba< 0.0001)。注射MPTP诱导多巴胺耗竭的MPTP药物+没锻炼组多巴胺(48.0±8.4 ng /毫克蛋白)与生理盐水+没锻炼组多巴胺(269.5±24.9 ng / ng蛋白质),对应于82%损耗。注射的MPTP药物诱导注射多巴胺耗竭的MPTP药物+运动组(4.9±0.2 ng多巴胺/毫克蛋白)与生理盐水+运动组(285.1±30.3 ng多巴胺/ ng蛋白质),对应于98%损耗。纹状体多巴胺水平没有明显差异之间的运动,没有运动动物注射的MPTP药物——或者saline-treated动物。然而,注射显著减少周转率MPTP药物+注射运动与MPTP药物+没锻炼组(gydF4y2BaFgydF4y2Ba(16)gydF4y2Ba= 8.30;gydF4y2BapgydF4y2Ba< 0.02)gydF4y2Ba
高效液相色谱分析,纹状体多巴胺及其代谢产物gydF4y2Ba

高效液相色谱分析,纹状体多巴胺和多巴胺水平营业额。这个图中显示的数据gydF4y2Ba表1gydF4y2Ba纹状体多巴胺的水平和周转率都5 d的运动(对应于10 d postlesioning),并在28 d运动(对应于42 d postlesioning)。在第五天的锻炼,有82 - 98%损耗的多巴胺在所有MPTP-treated动物(运动和不运动)与各自的生理盐水组(星号代表意义gydF4y2BapgydF4y2Ba< 0.0001)。纹状体多巴胺水平没有明显差异之间的运动,没有运动动物注射的MPTP药物——或者saline-treated动物。周转率显著升高所有MPTP-compared saline-treated动物(星号代表意义gydF4y2BapgydF4y2Ba< 0.0001)。有显著减少注射的周转率MPTP药物+注射运动与MPTP药物+没锻炼动物(哈希马克代表意义gydF4y2BapgydF4y2Ba< 0.02)。在28天的锻炼,> 68 - 78%损耗的纹状体多巴胺水平在所有MPTP-treated动物(运动和不运动)与各自的生理盐水组(星号代表意义gydF4y2BapgydF4y2Ba< 0.0001)。有显著增加纹状体多巴胺水平在生理盐水+运动与生理盐水+没锻炼组只(十字架代表意义gydF4y2BapgydF4y2Ba= 0.015)。锻炼对周转率没有影响。误差线表明SEM。gydF4y2Ba
纹状体多巴胺的高效液相色谱分析完成28 d的跑步机上跑步注射显示有显著影响的MPTP药物多巴胺耗竭程度(gydF4y2BaFgydF4y2Ba(16)gydF4y2Ba= 229.3;gydF4y2BapgydF4y2Ba< 0.0001)。注射MPTP诱导多巴胺耗竭的MPTP药物+没锻炼组多巴胺(77.9±12.0 ng /毫克蛋白)与生理盐水+没锻炼组多巴胺(246.9±19.8 ng / ng蛋白质),对应于68%损耗。注射的MPTP药物诱导注射多巴胺耗竭的MPTP药物+运动组(69.8±11.7 ng多巴胺/毫克蛋白)与生理盐水+运动组(315.2±9.0 ng多巴胺/ ng蛋白质),对应于78%损耗。此外,有一个重要的损伤状态之间的相互作用和运动对纹状体多巴胺水平(gydF4y2BaFgydF4y2Ba(16)gydF4y2Ba= 7.78;gydF4y2BapgydF4y2Ba= 0.015)。这种交互是因为saline-treated组显著影响锻炼,生理盐水+运动的老鼠有一个更高层次的纹状体多巴胺与盐水的老鼠。纹状体的多巴胺的水平没有显著差异注射比较MPTP药物+运动注射了MPTP药物+没锻炼的老鼠。注射的MPTP药物或运动没有明显影响,或者这两个因素之间的相互作用在周转率,比率为0.36注射对MPTP药物,注射0.34 MPTP药物+运动,0.34 0.26盐水,盐水+运动组。gydF4y2Ba
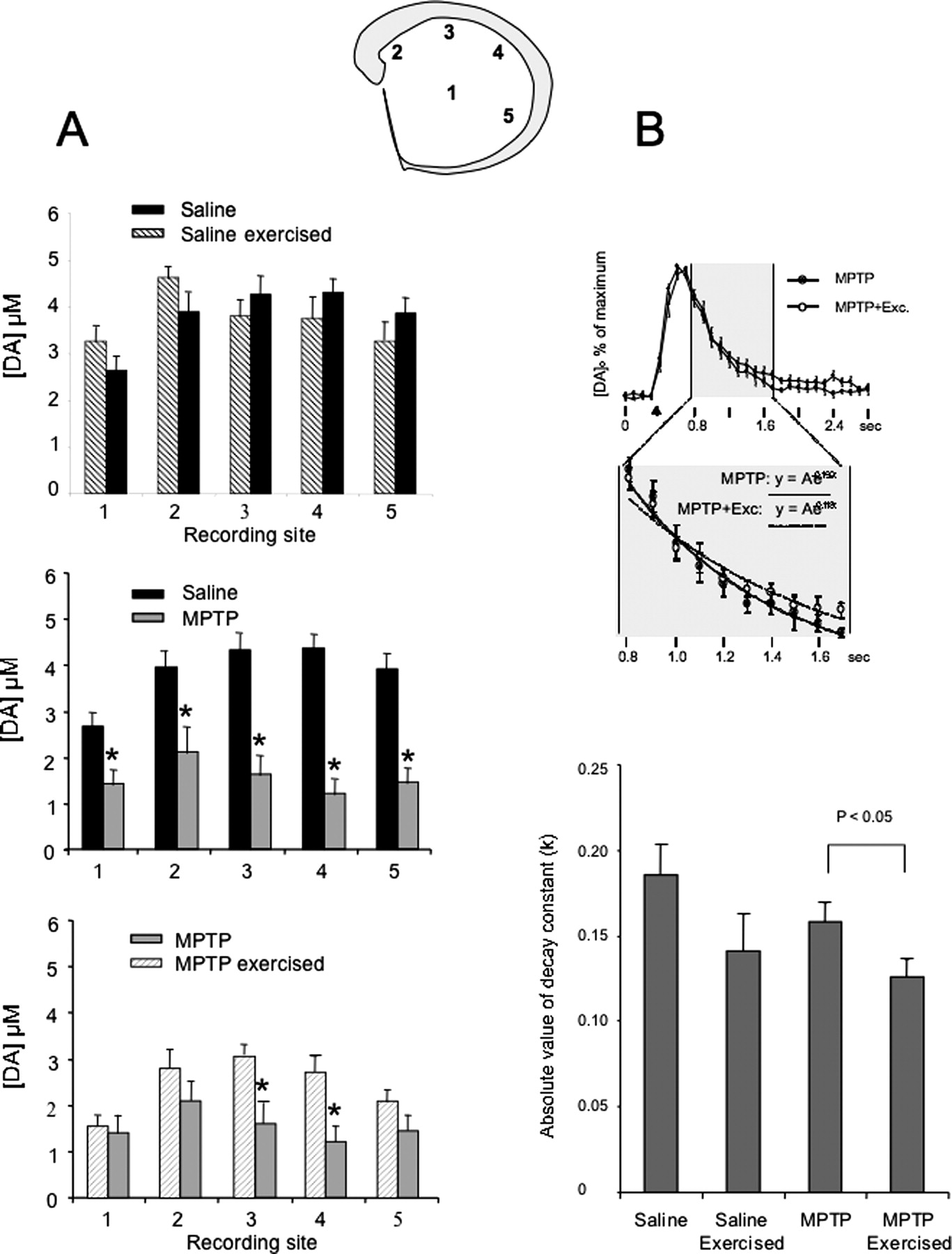
刺激多巴胺的释放引起intrastriatal决心使用快速扫描环绕伏安测量法完成28 d的跑步机上跑步。冠状切片的midstriatum采样与电极放置在5个站点,如所示gydF4y2Ba图4gydF4y2Ba。侧纹状体的测量,使用镜像位置。测量用四到六老鼠从每组和4片/鼠标。对于每一个动物,一个平均值确定在每个电极位置的平均收集的所有数据。电极刺激诱发多巴胺的释放随电极的位置(gydF4y2BaFgydF4y2Ba(20)gydF4y2Ba= 9.211;gydF4y2BapgydF4y2Ba< 0.001)在盐水和MPTP-treated老鼠。MPTP损伤显著降低电极刺激诱发多巴胺释放而saline-treated老鼠(gydF4y2BaFgydF4y2Ba(20)gydF4y2Ba= 26.450;gydF4y2BapgydF4y2Ba< 0.001)。锻炼的mptp缺陷型小鼠中有很大的影响依赖于电极位置(gydF4y2BaFgydF4y2Ba(20)gydF4y2Ba= 2.875;gydF4y2BapgydF4y2Ba= 0.039)。额外的分析使用gydF4y2Ba事后gydF4y2BatgydF4y2Ba测试表明,在电极位置有明显的多巴胺释放三个(gydF4y2BapgydF4y2Ba= 0.036)和4 (gydF4y2BapgydF4y2Ba= 0.023)但不是电极位置(gydF4y2BapgydF4y2Ba= 0.723),两个(gydF4y2BapgydF4y2Ba= 0.252)和5 (gydF4y2BapgydF4y2Ba= 0.141)注射在MPTP药物+运动小鼠注射了MPTP药物加上不运动的老鼠相比。锻炼没有unlesioned动物产生影响(没有整体效果,gydF4y2BaFgydF4y2Ba(20)gydF4y2Ba= 0.084,gydF4y2BapgydF4y2Ba= 0.781;没有锻炼和纹状体位置之间的相互作用,gydF4y2BaFgydF4y2Ba(20)gydF4y2Ba= 2.586,gydF4y2BapgydF4y2Ba= 0.088),但它确实影响损伤动物,根据纹状体中的位置(有一个互动的运动和位置)。我们intrastriatal刺激引起的多巴胺释放的机制被按顺序测试调查诱发多巴胺反应控制aCSF(包含2.4米gydF4y2Ba米gydF4y2BaCagydF4y2Ba2 +gydF4y2Ba和1.3gydF4y2Ba米gydF4y2Ba毫克gydF4y2Ba2 +gydF4y2Ba),减少钙gydF4y2Ba2 +gydF4y2BaaCSF(包含0.5米gydF4y2Ba米gydF4y2BaCagydF4y2Ba2 +gydF4y2Ba和3.2gydF4y2Ba米gydF4y2Ba毫克gydF4y2Ba2 +gydF4y2Ba),与正常aCSF冲刷之后,添加aCSF包含1μgydF4y2Ba米gydF4y2BaTTX。减少钙gydF4y2Ba2 +gydF4y2BaaCSF诱发多巴胺反应下降到25.85±5.44% (gydF4y2BangydF4y2Ba= 3)的控制,它返回到正常aCSF控制振幅与冲刷。后续添加TTX完全封锁了诱发多巴胺释放(gydF4y2BangydF4y2Ba= 3)。gydF4y2Ba

使用快速扫描环绕伏安测量法分析多巴胺的释放。的多巴胺释放量是决定在五个地区内纹状体(前囱水平1.00∼0.80前囱)包括(1)mid-striatum dorsomedial(2),(3)背,背外侧(4),(5)腹外侧代表老鼠从所有四组(见脑片插入)。gydF4y2Ba一个gydF4y2Ba,比较峰值由单一intrastriatal刺激多巴胺的释放(200 a, 0.1 ms)。,数据比较生理盐水和生理盐水+运动的老鼠。没有显著差异在任何地区的多巴胺的释放。中间,数据比较注射生理盐水和MPTP药物组。有显著减少小鼠注射多巴胺释放的MPTP药物和星号代表与生理盐水相比显著差异(gydF4y2BapgydF4y2Ba< 0.001)。底,注射数据比较MPTP药物注射和MPTP药物+运动的老鼠。有一个显著增加注射多巴胺释放的MPTP药物+运动小鼠背网站3和4小鼠注射了MPTP药物相比,和星号代表显著差异(gydF4y2BapgydF4y2Ba< 0.05)。gydF4y2BaBgydF4y2Ba、比较的多巴胺信号由intrastriatal刺激引发(峰值和衰变)。,一块平均时间和衰减的多巴胺信号峰值在电极位置注射4 MPTP药物+没有锻炼(gydF4y2BangydF4y2Ba= 7,注射填充圈)和MPTP药物+锻炼(gydF4y2BangydF4y2Ba= 11,开放的圆圈)。intrastriatal刺激当时交付的三角形(介于0和0.8 s)。数据归一化到peak-evoked多巴胺信号。中间的面板显示了衰减阶段的图表(阴影部分),这是符合一个指数函数。最适合注射平均数据从MPTP药物+没有运动老鼠注射(实线)和MPTP药物+运动小鼠(虚线)所示。底部面板显示了平均衰减常数(gydF4y2BakgydF4y2Ba)获得的指数衰减阶段的每个记录和显示每组(均值±SEM)。MPTP +运动小鼠注射了MPTP药物相比显著降低衰减常数加上不运动的老鼠(gydF4y2BapgydF4y2Ba< 0.05;gydF4y2BatgydF4y2Ba测试)。类似的趋势的盐水+运动较生理盐水+没锻炼动物(gydF4y2BapgydF4y2Ba= 0.13;gydF4y2BatgydF4y2Ba测试)。Exc,锻炼。gydF4y2Ba
多巴胺信号诱发的衰变率intrastriatal刺激也比较网站4之间的每个组(见材料与方法确定衰变速率常数,−gydF4y2BakgydF4y2Ba)。网站4被选中,因为这个网站显示最大的运动不同的多巴胺释放峰值mptp缺陷型组。运动有影响的衰减常数(−诱发多巴胺信号gydF4y2BakgydF4y2Ba)。gydF4y2Ba事后gydF4y2Ba分析显示显著放缓注射多巴胺的衰变率MPTP药物+运动组(gydF4y2BangydF4y2Ba= 11)单独注射相比MPTP药物组(gydF4y2BangydF4y2Ba= 7;gydF4y2BapgydF4y2Ba< 0.05;gydF4y2BatgydF4y2Ba测试)。生理盐水+运动组之间也出现了类似的趋势(gydF4y2BangydF4y2Ba= 11)和生理盐水组(gydF4y2BangydF4y2Ba= 11;gydF4y2BapgydF4y2Ba= 0.13)。gydF4y2Ba
正如先前所显示的,有一个显著减少纹状体注射DAT和TH蛋白表达MPTP药物+运动小鼠注射了MPTP药物老鼠相比(gydF4y2BapgydF4y2Ba< 0.05)(gydF4y2Ba费舍尔et al ., 2004gydF4y2Ba)。MPTP-treated老鼠也显示有显著减少TH和DAT蛋白表达与saline-treated老鼠(gydF4y2BapgydF4y2Ba< 0.001)。在TH没有显著差异,DAT蛋白质之间的观察生理盐水+运动和生理盐水的老鼠。这些数据所示gydF4y2Ba图5gydF4y2Ba。确定蛋白质水平的变化是伴随着变化的相对表达TH或DAT mRNA转录在中脑多巴胺能神经元,gydF4y2Ba原位gydF4y2Ba杂交组织化学结合粒计算的emulsion-dipped部分使用。从所有四组老鼠进行了分析后28 d锻炼和这些数据所示gydF4y2Ba图6gydF4y2Ba。MPTP损伤引起显著减少的相对表达TH和DAT转录表达在中脑多巴胺神经元(gydF4y2BaFgydF4y2Ba(12)gydF4y2Ba= 26.1,gydF4y2BapgydF4y2Ba< 0.001和gydF4y2BaFgydF4y2Ba(12)gydF4y2Ba= 37.29,gydF4y2BapgydF4y2Ba分别为< 0.001)。运动引起显著减少TH和DAT mRNA转录表达在MPTP-treated saline-treated但不是老鼠。具体来说,尽管锻炼对TH成绩单没有整体效应表达式(gydF4y2BaFgydF4y2Ba(12)gydF4y2Ba= 2.26;gydF4y2BapgydF4y2Ba= 0.133),有显著注射运动和MPTP药物之间的交互损伤(gydF4y2BaFgydF4y2Ba(12)gydF4y2Ba= 51.7;gydF4y2BapgydF4y2Ba< 0.001)。这种交互是因为TH mRNA表达减少28%的生理盐水+运动组与生理盐水+没有运动组。锻炼也导致DAT转录表达明显降低,(gydF4y2BaFgydF4y2Ba(12)gydF4y2Ba= 73.2;gydF4y2BapgydF4y2Ba< 0.001),有显著注射运动和MPTP药物之间的交互损伤(gydF4y2BaFgydF4y2Ba(12)gydF4y2Ba= 33.7;gydF4y2BapgydF4y2Ba< 0.001)。这种交互又因为DAT mRNA表达减少43%的生理盐水+运动组较生理盐水+没有运动组。gydF4y2Ba

纹状体TH和DAT蛋白质的分析。gydF4y2Ba一个gydF4y2Ba,gydF4y2BaBgydF4y2Ba纹状体的相对表达TH (gydF4y2Ba一个gydF4y2Ba)和DAT (gydF4y2BaBgydF4y2Ba)确定使用免疫组织化学和计算机辅助图像分析后28 d的运动。部分的midstriatum(≥6 - 8部分从4个不同的老鼠在每个组)和抗体对TH或DAT应用。部分进行扫描和相对光密度意味着+ SEM为每个组决定。MPTP导致大幅下降的纹状体TH和DAT蛋白质的表达水平与saline-treated老鼠相比,和星号代表意义gydF4y2BapgydF4y2Ba< 0.001。注射之间存在显著的交互运动和MPTP药物,注射的明显降低MPTP药物+注射运动与MPTP药物+不运动的老鼠,和哈希马克代表意义gydF4y2BapgydF4y2Ba< 0.05。gydF4y2Ba

分析TH和DAT mRNA在中脑黑神经元。的相对表达TH和DAT mRNA在中脑多巴胺神经元使用决定gydF4y2Ba原位gydF4y2Ba杂交组织化学颗粒计数emulsion-dipped部分紧随其后。代表部分显示决心TH mRNA表达的神经元所示面板gydF4y2Ba一个gydF4y2Ba(生理盐水),gydF4y2BaBgydF4y2Ba(生理盐水+运动),gydF4y2BaCgydF4y2Ba注射(MPTP药物)gydF4y2BaDgydF4y2Ba注射(MPTP药物+运动)。各自的分析数据的TH或DAT下图所示每组图像。至少有120个神经元,谷物被计算为每个治疗组。MPTP损伤导致明显降低TH和DAT成绩单与saline-treated组相比,和星号代表意义gydF4y2BapgydF4y2Ba< 0.001。统计之间的差异被盐水,盐水+运动团体TH和DAT成绩单,和十字架代表意义gydF4y2BapgydF4y2Ba< 0.001。没有明显差异注射之间的转录表达MPTP药物注射和MPTP药物+运动组。代表部分显示神经元的DAT mRNA表达所示gydF4y2BaEgydF4y2Ba(生理盐水),gydF4y2BaFgydF4y2Ba(生理盐水+运动),gydF4y2BaGgydF4y2Ba注射(MPTP药物)gydF4y2BaHgydF4y2Ba注射(MPTP药物+运动)。误差线表明SEM。gydF4y2Ba
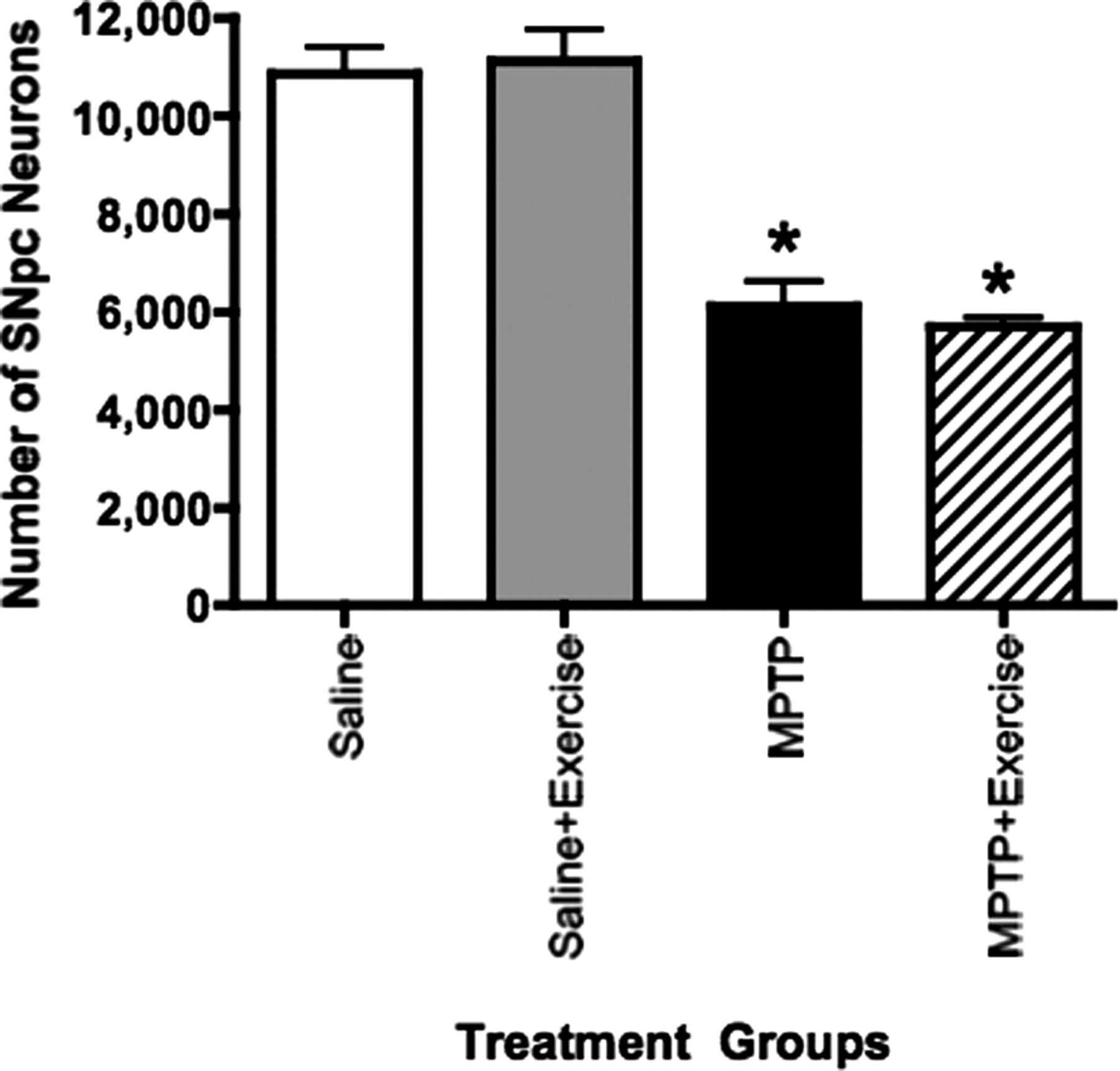
作为衡量中脑多巴胺能神经元变性黑的完整性,我们确定的数量TH-ir SNpc使用无偏stereological计数小鼠神经元的生理盐水,盐水+运动,注射MPTP和MPTP药物+运动组(gydF4y2BangydF4y2Ba每组)= 3 (gydF4y2Ba图7gydF4y2Ba)。MPTP损伤导致的数量明显降低TH-ir SNpc神经元(gydF4y2BaFgydF4y2Ba(8)gydF4y2Ba= 106.9;gydF4y2BapgydF4y2Ba< 0.001)。然而,运动造成的数量无显著影响TH-ir SNpc神经元(gydF4y2BaFgydF4y2Ba(8)gydF4y2Ba= 0.022;gydF4y2BapgydF4y2Ba= 0.885),注射之间没有显著的交互运动和MPTP药物TH-positive神经元的数量。gydF4y2Ba

SNpc测定多巴胺能细胞数量。无偏stereological TH-ir神经元在老鼠身上进行计数(gydF4y2BangydF4y2Ba从所有四组每组= 3)结束时28 d运行方案。MPTP损伤导致了∼50%下降SNpc多巴胺能神经元。之间没有统计上的显著差异盐水,盐水+运动老鼠注射以及没有区别MPTP药物注射和MPTP药物+运动组,表明没有效果的运动SNpc多巴胺能神经元的数量。星号代表从盐水组显著差异(gydF4y2BapgydF4y2Ba< 0.0001)。gydF4y2Ba
讨论gydF4y2Ba
此前报道,密集的跑步机锻炼导致改善电机性能的mptp缺陷型(gydF4y2Ba蒂勒森et al ., 2003gydF4y2Ba;gydF4y2Ba费舍尔et al ., 2004gydF4y2Ba)和saline-treated老鼠(gydF4y2Ba费舍尔et al ., 2004gydF4y2Ba)。在目前的研究中,我们也证明了密集的跑步机锻炼导致延时增加下降加速rotarod(改善)。这一发现表明,密集的跑步机锻炼可能通过自适应变化的基底神经节和电机电路,导致相关改善电机的任务(平衡)注射在MPTP药物和saline-treated动物。然而,我们的研究结果表明,锻炼的有利影响是伴随着微分对多巴胺能系统的影响可能是依赖黑损伤的存在与否(损伤与unlesioned)。具体来说,虽然在两组剧烈运动改善发动机性能,利用高效液相色谱分析,我们发现这个功能效益在总伴随着显著增加纹状体多巴胺后28 d运动生理盐水+运动与生理盐水+不运动的老鼠。没有明显差异总共注射纹状体多巴胺水平MPTP药物老鼠,检查5或后28 d的跑步机上跑步,而没有相应的运动组。此外,运动没有影响多巴胺的时间进程返回的mptp缺陷型小鼠中。具体来说,我们观察到在所有的mptp缺陷型小鼠纹状体多巴胺的部分返回至28天与第五天相比,这效果观察到相同的程度在运动和不运动的老鼠,和依照先前的报道(gydF4y2BaRicaurte et al ., 1986gydF4y2Ba;gydF4y2BaBezard et al ., 2000gydF4y2Ba;gydF4y2BaJakowec et al ., 2004gydF4y2Ba)。虽然总组织儿茶酚胺的高效液相色谱分析是一个很好的衡量总多巴胺的池,包括突触,extra-synaptic,水泡,和胞质,它可能不是一个准确的估计的多巴胺释放量的活动(gydF4y2Ba加里et al ., 1997gydF4y2Ba;gydF4y2BaYavich麦克唐纳,2000gydF4y2Ba;gydF4y2BaDentresangle et al ., 2001gydF4y2Ba)。gydF4y2Ba
与总组织高效液相色谱分析,使用快速扫描环绕伏安测量法在纹状体片老鼠28 d,我们观察到一个注射stimulus-evoked多巴胺释放增加exercise-dependent MPTP药物+运动相比,注射了MPTP药物+没锻炼的老鼠。我们的研究结果表明,stimulus-evoked多巴胺释放是水泡,因为它减少了细胞外钙减少gydF4y2Ba2 +gydF4y2Ba这是完全被堵塞的NagydF4y2Ba+gydF4y2Ba渠道TTX先前报道(gydF4y2Ba克拉格和绿地,1997gydF4y2Ba;gydF4y2Ba陈和大米,2001年gydF4y2Ba)。exercise-dependent效果未见在生理盐水+运动与生理盐水+没锻炼的老鼠。有趣的是,运动影响stimulus-evoked注射多巴胺释放的MPTP药物+练习鼠标背外侧纹状体中最为明显。一个可能的解释对于这个练习和地区的特定效应可能是由于use-dependent形式的突触可塑性。高度的参与要求背外侧纹状体的前肢和后肢运动在跑步机上运动支持研究表明选择性增加血流和代谢活动在背外侧纹状体在跑步机上锻炼(gydF4y2BaCospito Kultas-Ilinsky, 1981gydF4y2Ba;gydF4y2BaEbrahimi et al ., 1992gydF4y2Ba;gydF4y2Ba布朗和锋利,1995年gydF4y2Ba;gydF4y2BaHolschneider et al ., 2003gydF4y2Ba;gydF4y2Ba阮et al ., 2004gydF4y2Ba)。gydF4y2Ba
快速扫描环绕伏安测量法是一种技术,允许精确的解剖定位和检查诱发多巴胺释放许多不同的地点在同一个大脑切片在一个录音,从而提供检测多巴胺释放中的地区差异的能力。虽然一个优势gydF4y2Ba在活的有机体内gydF4y2Ba记录的保留完整的传入通路,刺激的内侧前脑束(地铁消防队)可能是变量在这些传入纤维和导致多巴胺释放的差异即使在一百微米之间的纹状体记录网站(gydF4y2Ba天et al ., 1989gydF4y2Ba)。脑片可能不保留一个完整的黑途径但它避免诱发释放的变化,因为整个记录字段是兴奋和提供了一个合理的工具研究这种神经递质系统响应(gydF4y2Ba帕特尔和大米,2006年gydF4y2Ba)。在一组控制实验中,我们证实,多巴胺的释放实际上是介导通过动作电位进行快速扫描环绕伏安测量法研究存在的钠离子通道阻滞剂TTX。gydF4y2Ba
在协议与我们的研究中,补充多巴胺释放的变化已报告受伤后使用6-OHDA多巴胺能系统损伤和PD患者在运动锻炼(gydF4y2BaZhang et al ., 1988gydF4y2Ba;gydF4y2Ba加里et al ., 1997gydF4y2Ba;gydF4y2Ba大内et al ., 2001gydF4y2Ba)。此外,最近的一项研究gydF4y2BaO 'Dell et al。(2006)gydF4y2Ba使用6-OHDA纹状体损伤大鼠,支持我们的研究结果,改善电机性能未必是伴随着总纹状体多巴胺水平运动后的变化。具体地说,这组报道保持更高回报的运动行为没有纹状体多巴胺水平的回归prelesioning基线(gydF4y2BaO 'Dell et al ., 2006gydF4y2Ba)。我们的研究和报道gydF4y2BaO 'Dell et al。(2006)gydF4y2Ba相比其他显示部分返回纹状体多巴胺neurotoxicant-lesioned啮齿动物模型中进行锻炼(gydF4y2Ba蒂勒森et al ., 2003gydF4y2Ba)。一个可能解释这种差异的差异可能是因为MPTP-lesioning养生和运动模式。在我们的研究中,我们使用了一个MPTP-lesioning时间表,导致中等程度的黑多巴胺能细胞损失和启动注射后锻炼5 d MPTP药物损伤,细胞死亡时完成。相反,研究表明多巴胺部分返回使用温和的损伤方案,立即启动跑步机跑步损伤后(24小时内),当潜在的DAT的差别运动对这些基因可能会影响神经毒素吸收的数量和程度的损伤(gydF4y2BaGainetdinov et al ., 1997gydF4y2Ba)。无偏stereological计数中脑多巴胺能神经元的DAT的差别在我们的研究表明,运动对这些独立的幸存的物质的数量nigral神经元。gydF4y2Ba
使用免疫组织化学分析,我们发现28 d剧烈运动的差别导致了对这些DAT(一种蛋白质负责吸收和清除的多巴胺细胞外空间)和TH(酶负责多巴胺生物合成的病原反应步骤)(gydF4y2BaJackson-Lewis et al ., 1995gydF4y2Ba;gydF4y2Ba费舍尔et al ., 2004gydF4y2Ba)的蛋白质。这个发现支持我们以前的工作证明抑制TH和DAT注射蛋白质MPTP药物+运动小鼠使用西方免疫印迹和免疫组织化学染色法,两种互补的技术研究蛋白质表达水平和解剖模式分布(gydF4y2Ba伯克et al ., 1990gydF4y2Ba;gydF4y2BaJakowec et al ., 2004gydF4y2Ba)。来验证这种减少的纹状体DAT和TH蛋白质并不是因为运动型黑细胞死亡,中脑多巴胺神经元的数量基于TH免疫反应性测定使用无偏stereological计数技术。比较运动,没有运动组没有显著差异的总数中脑多巴胺神经元之间注射要么MPTP药物注射和MPTP药物+运动或生理盐水,盐水+运动的老鼠,这表明没有额外的运动引起的细胞死亡。这一发现表明,DAT和TH蛋白表达下调的运动在注射后幸存的中脑多巴胺神经元MPTP药物损伤。Downregulation DAT可能导致增加突触多巴胺能系统的可用性和代表一个补偿效应,因为受伤的运动状态。事实上,使用快速扫描环绕伏安测量法分析的衰变率在大脑纹状体多巴胺信号由一个刺激诱发片,我们观察到注射多巴胺衰变速率较慢的MPTP药物+注射运动与MPTP药物+不运动的老鼠,可能与DAT的下降有关。多巴胺释放的增加在行使老鼠也可以反射更多的扩散的多巴胺突触远离记录网站当DAT降低(gydF4y2Ba克拉格和大米,2004年gydF4y2Ba)。运动诱发多巴胺释放增加DAT-related延长一起背外侧纹状体的多巴胺信号可能构成一个机制剧烈运动促进正常电机电路因为层次的突触可塑性,有限公司和LTP,由多巴胺调制。在背外侧纹状体,衰变的多巴胺释放增加,减少可能发挥重要作用在维护正常的有限公司(gydF4y2Ba卡拉布雷西et al ., 1992gydF4y2Ba;gydF4y2Ba史密斯et al ., 2001gydF4y2Ba;gydF4y2BaAkopian和沃尔什,2007年gydF4y2Ba)。gydF4y2Ba
DAT和TH蛋白的表达,以及纹状体多巴胺的水平,是密切相关的。例如,DAT淘汰赛老鼠已经被证明表达减少纹状体TH蛋白(gydF4y2Ba贾比尔et al ., 1999gydF4y2Ba)。尽管准确DAT和TH之间的联系还不清楚,这是推测多巴胺突触生物利用度的增加,DAT的差别通过对这些基因可能导致增加内多巴胺受体的激活,TH(差别导致对这些gydF4y2BaFauchey et al ., 2000gydF4y2Ba)。支持我们的研究结果,研究与神经影像学显示DAT PD患者接受汽车差别迅速对这些活动(步行)(gydF4y2Ba大内et al ., 2001gydF4y2Ba)。DAT的差别类似的对这些活动可以诱导在年轻小鼠暴露于环境浓缩,这可以证明神经注射对MPTP药物损伤(gydF4y2BaBezard et al ., 2003gydF4y2Ba)。gydF4y2Ba
在我们的研究中,我们使用gydF4y2Ba原位gydF4y2Ba杂交组织化学检查的mRNA转录水平的表达TH和DAT幸存的黑多巴胺能神经元是否改变转录可以解释运动性抑制纹状体蛋白质。我们发现减少的表达TH和DAT mRNA在生理盐水+运动相比,注射生理盐水的老鼠而不是MPTP药物+运动小鼠注射了MPTP药物相比。运动对mRNA转录表达的影响可能依赖于前一个受伤的存在黑多巴胺能神经元。MPTP可导致减少在mRNA表达幸存的中脑神经元,这可能潜在影响的运动(gydF4y2BaJakowec et al . 2004gydF4y2Ba)。因为运动给纹状体改变蛋白质的转录表达的变化不能占存活小鼠注射多巴胺能神经元变性黑MPTP药物,运动可能通过其他机制影响TH和DAT的翻译,轴突易位或营业额。gydF4y2Ba
总之,我们的结果表明,密集的跑步机锻炼导致注射改善电机性能的MPTP药物和盐水的老鼠,这行为改善也观察到在一个相关的运动任务。锻炼的有益的影响可能是由于多巴胺能神经传递的改变,这可能是正常和受伤的基底神经节之间的不同。运动导致补偿性的mptp缺陷型小鼠的变化导致增加突触释放多巴胺通过增加可用性,减少吸收,减少腐烂。在盐水的老鼠,运动可能通过影响多巴胺水平升高,因为增加的生物合成通过增加活动。目前,连接运动和多巴胺能神经传递的分子机制是未知的。然而,文献报道表明神经营养因子的作用,如脑源性神经营养因子、纤维母细胞生长因子,或glial-derived神经营养因子,通过供应下游通路的激活,如蛋白质激酶,可能影响突触可塑性和终端神经传递(gydF4y2Bagomez - pinilla et al ., 1997gydF4y2Ba,gydF4y2Ba2002年gydF4y2Ba;gydF4y2Ba科恩et al ., 2003gydF4y2Ba)。glutamatergic层次的途径可能是另一个候选人系统参与了多巴胺释放的状况改变,因为它是一个重要的调制器在纹状体多巴胺的释放,和我们之前的工作表明改变密度的纹状体注射谷氨酸的immunolabeling MPTP药物+运动小鼠与久坐的MPTP缺陷型小鼠相比gydF4y2BaMolteni et al ., 2002gydF4y2Ba;gydF4y2Ba费舍尔et al ., 2004gydF4y2Ba;gydF4y2Ba迪特里希et al ., 2005gydF4y2Ba)。整体而言,这些结果表明,经济复苏的运动行为可以通过小说实际上发生在基底神经节和补偿机制,这种复苏的机制可能不同的损伤比nonlesioned基底神经节。这些发现可能是重要的治疗基底神经节的神经退行性疾病,包括帕金森病、增强的神经可塑性通过锻炼可能会导致改变多巴胺可用性(释放和吸收),这反过来可能会发挥更重要作用在维护正常的突触连接比恢复绝对的多巴胺的水平。gydF4y2Ba
脚注gydF4y2Ba
这项工作是帕金森病基金会支持的团队帕金森洛杉矶,惠蒂尔帕金森病教育集团,国家神经疾病和中风研究所授予RO1 NS44327-1,和美国军队的神经毒素暴露治疗研究项目拨款w81xwh - 04 - 1 - 0444。特别感谢南加州大学的朋友帕金森病研究小组,包括乔治和玛丽卢布恩和沃尔特和苏珊Doniger。我们感谢玛格丽特·e .大米和查尔斯Meshul他们深刻的讨论研究,特别感谢Nicolaus帕斯卡,多米尼克的支持。gydF4y2Ba
- 信件应该写给迈克尔·w·Jakowec博士gydF4y2Ba部的神经学、南加州大学、圣巴首页勃罗街1333号,mca - 241, 90033年洛杉矶CA。gydF4y2Bajakowec在}{surgery.usc.edugydF4y2Ba
引用gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba
- ↵gydF4y2Ba








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}